Ориентировочный рефлекс как основа внимания
Активирующая ретикулярная система с ее восходящими и нисходящими волокнами является нейрофизиологическим аппаратом, обеспечивающим одну из наиболее важных форм рефлекторной деятельности, известную под названием ориентировочного (или ориентировочно-исследовательского) рефлекса.
Его значение для понимания физиологических основ внимания настолько велико, что на нем следует остановиться специально.
Каждый безусловный рефлекс, имеющий в своей основе какое-либо биологически важное для животного воздействие (пищевое, болевое, половое), вызывает избирательную систему ответов на эти раздражители с одновременным торможением всех реакций на побочные. Такой же избирательный характер носят и условные рефлексы. При них одна система реакций, подкрепляемая безусловным раздражителем, доминирует, в то время как все остальные побочные реакции тормозятся. Можно сказать, что как безусловные, так и сформированные на их основе условные рефлексы создают известный доминирующий очаг возбуждения, протекание которого подчиняется доминанте.
Среди всех видов рефлекторной деятельности нужно, однако, выделить один, при котором поведение животного не возбуждается ни одним из перечисленных выше мотивов поведения и который не является ни пищевым, ни оборонительным, ни половым рефлексом. Основой этой деятельности является активная реакция животного на каждое изменение обстановки, которое и вызывает у животного общее оживление и ряд избирательных реакций, направленных на ознакомление с этими изменениями в ситуации. И. П. Павлов назвал этот вид рефлексов «ориентировочными рефлексами«, или «рефлексами «что такое?»».
Ориентировочный рефлекс выражается в ряде отчетливых электрофизиологических, сосудистых и двигательных реакций, появляющихся каждый раз, когда в обстановке, окружающей животное, возникает что-нибудь необычное или существенное. К этим реакциям относятся :
- поворот глаз и головы в сторону нового объекта;
- реакция настораживания или прислушивания.
А у человека — появление кожно-гальванической реакции (изменение сопротивляемости кожи к электрическому току или появление собственных электрических потенциалов кожи), сосудистые реакции (сужение сосудов руки с расширением сосудов головы), изменение дыхания, наконец, возникновение явлений «десинхронизации» в биоэлектрических реакциях мозга, выражающихся в депрессии «альфа ритма» (электрических колебаний 10-12 в сек, характерных для работы мозговой коры в спокойном состоянии). Все эти явления можно наблюдать каждый раз, когда возникает реакция настораживания, или ориентировочный рефлекс, вызываемый появлением нового или существенного для субъекта раздражителя.
Среди ученых нет еще единодушного ответа на вопрос, является ли ориентировочный рефлекс безусловной или условной реакцией.
По своему врожденному характеру ориентировочный рефлекс можно отнести к числу безусловных. Животное отвечает реакцией настораживания на любые новые или существенные раздражители без всякого обучения; по этому признаку ориентировочный рефлекс относится к числу безусловных, врожденных реакций организма. Наличие особых нейронов, которые отвечают разрядами на каждое изменение ситуации, указывает на то, что в его основе лежит действие определенных нервных приборов.
С другой стороны, ориентировочный рефлекс обнаруживает ряд признаков, существенно отличающих его от обычных безусловных рефлексов: при неоднократном повторении одного и того же раздражителя явления ориентировочного рефлекса скоро угасают, организм привыкает к этому раздражителю, и его предъявления перестают вызывать описанные реакции. Это исчезновение ориентировочных реакций на повторяющиеся раздражители называется привыканием (habituation).
Следует отметить, что это исчезновение ориентировочного рефлекса по мере привыкания может быть временным явлением, и достаточно небольшого изменения в раздражителе, чтобы ориентировочная реакция снова возникла. Это явление возникновения ориентировочного рефлекса при малейшем изменении раздражения иногда называется реакцией «пробуждения» (или arousal). Характерно, что такое появление ориентировочного рефлекса, как мы уже отмечали выше, может иметь место не только при усилении, но и при ослаблении привычного раздражителя и даже при его полном исчезновении. Так, достаточно сначала «угасить» ориентировочные рефлексы на ритмически предъявляемые раздражители, а затем, после того как ориентировочные реакции на каждое раздражение угасли в результате привыкания, пропустить один из ритмически предъявляемых раздражителей. В этом случае отсутствие ожидаемого раздражителя вызовет появление ориентировочного рефлекса.
Всеми этими признаками своей динамики ориентировочный рефлекс существенно отличается от безусловного рефлекса. Следует отметить и тот факт, что ориентировочный рефлекс может быть вызван и условным раздражителем: его можно получить, если предъявлять животному условный сигнал, который будет говорить о появлении какого-либо изменения в окружающей обстановке. У человека таким сигналом может быть слово, которое легко вызывает у него явления подготовки, настораживания, ожидания появления сигнала и т. п.
Было бы неверным думать, что ориентировочный рефлекс носит характер общей, генерализированной активации организма. На самом деле он может иметь дифференцированный, избирательный характер, причем эта избирательность может проявляться как по отношению к возникающим сигналам, так и по характеру той готовности эффекторных двигательных аппаратов, которые вызываются «настороженностью».
Это легко видеть, если длительно предъявлять испытуемому какой-нибудь один сигнал, например звук определенной высоты, тогда в силу привыкания все реакции на этот звук будут угашены, однако это «привыкание» будет носить избирательный характер, и стоит только минимально изменить высоту звука, чтобы весь комплекс ориентировочных реакций снова появился. Такой прием позволил советскому исследователю Е. Н. Соколову объективно оценить ту избирательность, которой характеризуются ориентировочные реакции («реакции пробуждения») в отношении дифференцированных сигналов, и говорить о «нервной модели стимула», которая обнаруживается с помощью этого приема.
Лурия А. Р. Лекции по общей психологии. — СПб.: Питер, 2004. — 320 с. С. 176-178.
ориентировочный рефлекс — это… Что такое ориентировочный рефлекс?
- ориентировочный рефлекс
- — сложная реакция животных и человека на новизну стимула, названная И. П. Павловым рефлексом «что такое?». Биологический смысл О. р. — создание условий для лучшего восприятия раздражителя. Это достигается за счет появления комплекса соматических, вегетативных реакций и изменения уровня активации центральной нервной системы при общем торможении или нарушении текущей деятельности организма. Выделяют три основные группы компонентов О. р.:
1) двигательные;
2) вегетативные;
3) изменение уровня активации центральной нервной системы. К двигательным компонентам О. р. относятся:
1) реакции, обеспечивающие поворот глаз в направлении зрительного раздражителя, настораживание ушей, принюхивание;
2) реакции, обеспечивающие поддержание позы и приближение к стимулу.
К вегетативным компонентам О. р. относятся сосудистые реакции в виде сужения сосудов конечностей и расширения сосудов головы, обеспечивающие увеличение кровоснабжения мозга, кожно — галъваническую реакцию (КГР), изменение частоты сердцебиения и дыхания.
В качестве возбудителей О. р. помимо новизны стимула выделяют такие его свойства, как изменение стимула, его сложность, двусмысленность, удивительность. В том случае, когда стимул и след, оставленный ранее предъявленными раздражителями (см. нервная модель стимула), совпадают, О. р. не возникает. По мере повторения нового стимула, вызывающего О. р., происходит его ослабление, угасание. В зависимости от широты распространения возбуждения в нервной системе различают генерализованную и локальную формы О. р. В зависимости от длительности выделяют долго длящийся тонический и быстро завершающийся физический О. р. В процессе угасания генерализованный О. р. сменяется локальным, тонический физическим О. р.
Краткий психологический словарь. — Ростов-на-Дону: «ФЕНИКС».
Л.А.Карпенко, А.В.Петровский, М. Г. Ярошевский.
1998.
- ориентировочный рефлекс
- Этимология.
Происходит от лат. оrientis — восток и reflexus — отраженный.
Категория.
Форма рефлекса.
Структура.
Комплекс реакций организма в ответ на новизну раздражителя. К этим реакциям относятся: движение головы и глаз в направлении источника раздражения, расширение сосудов мозга при одновременном сужении периферических сосудов, изменение дыхания и электрического сопротивления кожи, возрастание тонуса мышц, повышение физиологической активности коры больших полушарий головного мозга (уменьшения амплитуды альфа-ритма), повышение чувствительности анализаторов и возрастание критической частоты слияния ощущений. При действии нового раздражителя появляются все компоненты ориентировочного рефлекса, образуя т. н. генерализованный ориентировочный рефлекс, который постепенно, после 15 — 20 предъявлений нового раздражителя, угасает, приобретая форму локального ориентировочного рефлекса. На основании сходства угасания ориентировочного рефлекса с угасанием условного рефлекса И.П.Павлов трактовал оба процесса как результат развития внутреннего торможения.
Психологический словарь.
И.М. Кондаков.
2000.
- ориентировочная реакция
- орудийные действия животных
Смотреть что такое «ориентировочный рефлекс» в других словарях:
Ориентировочный Рефлекс — комплекс реакций организма в ответ на новизну раздражителя, в который входят: движение головы и глаз в направлении источника раздражения, расширение сосудов мозга при одновременном сужении периферичес … Психологический словарь
ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС — (ориентировочная реакция) сложная реакция организма на новый раздражитель, направленная на мобилизацию систем организма для выработки соответствующего действия в новой ситуации. О. р. лежит в основе ориентировочной деятельности человека, т. е.… … Энциклопедический словарь по психологии и педагогике
Ориентировочный рефлекс — специфическая реакция животных, направленная на получение дополнительной информации путем создания условий для лучшего восприятия раздражителей. Выражается в общем прекращении текущей деятельности (стоп реакция), ориентации анализаторов на… … Словарь дрессировщика
ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС — См. ориентировочная реакция … Толковый словарь по психологии
рефлекс ориентировочный — комплекс реакций организма в ответ на новизну раздражителя сложная реакция животных и человека на новизну стимула, названная И. П. Павловым рефлексом «что такое?». Его биологический смысл создание условий для лучшего восприятия раздражителя. Это… … Большая психологическая энциклопедия
рефлекс безусловный — (рефлекс врожденный) наследственно закрепленная стереотипная форма реагирования на биологически значимые воздействия внешнего мира или на изменения внутренней среды организма. Рефлекс, всегда реализуемый при действии на организм определенных… … Большая психологическая энциклопедия
рефлекс врожденный — (рефлекс врожденный) наследственно закрепленная стереотипная форма реагирования на биологически значимые воздействия внешнего мира или на изменения внутренней среды организма. Рефлекс, всегда реализуемый при действии на организм определенных … Большая психологическая энциклопедия
РЕФЛЕКС ОРИЕНТИРОВОЧНЫЙ — сложная реакция животного или человека, вызываемая действием новых раздражителей или изменением старых, названная И.П. Павловым рефлексом «что та кое?». С Р. о. начинается любой сложнорефлекторный акт организма … Психомоторика: cловарь-справочник
Ориентировочная реакция — (рефлекс «Что такое?», по И. П. Павлову) комплекс сдвигов в разных системах организма животного или человека, вызываемый любым неожиданным изменением ситуации и обусловленный особой активностью центральной нервной системы. Изменения в… … Большая советская энциклопедия
ФОТОТАКСИС — Ориентировочный рефлекс на свет. Положительный фототаксис – движение по направлению к источнику света, отрицательный – от него. Этот термин предназначен для обозначения таких движений у животных; в отношении растений используется термин… … Толковый словарь по психологии
ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС — Студопедия
Ориентировочный рефлекс, или рефлекс «Что такое?» был открыт И.П. Павловым. Он описал его как комплекс двигательных реакций, который возникал на неожиданное появление нового стимула. Собака поворачивала голову, глаза, настораживала уши в направлении нового раздражителя. В работах Ю. Конорского [1970] этот комплекс соматических реакций получил название рефлекса прицеливания.
Рассматривая ориентировочный рефлекс как реакцию, направленную на восприятие нового стимула, И.П. Павлов обратил основное внимание на его тормозную, прерывающую функцию в отношении текущей условно-рефлекторной деятельности. Во время ориентировочной реакции условный рефлекс либо полностью подавлялся, либо его выполнение ухудшалось. Это свойство ориентировочной реакции нарушать текущую деятельность, в том числе и процесс обучения (выработку условного рефлекса), привело И.П. Павлова к мысли о необходимости построить исследовательский корпус в виде «Башни молчания». Это позволило бы исключить тормозные эффекты ориентировочного рефлекса (ОР) на формирование и исполнение условных рефлексов.
Хотя вначале ОР и представлялся И. П. Павлову и его коллегам преимущественно как внешний тормоз условного рефлекса, они описали многие его особенности, которые выходили за пределы тормозной функции. Так был открыт растормаживающий эффект ориентировочной реакции. Он состоял в том, что ранее выработанные, а затем угашенные условные связи под влиянием ОР вновь восстанавливались. Сегодня мы интерпретировали бы эти данные как способность восстанавливать подавленные связи в процедурной долговременной памяти. В лаборатории И.П. Павлова также были получены первые факты о влиянии ОР на формирование условного рефлекса. Во время процедуры выработки условной связи, а также при формировании ассоциации между двумя индифферентными раздражителями было отмечено усиление или появление ориентировочных реакций. В опытах И.О. Нарбутович и Н.А. Подкопаева [1936] с выработкой у собаки ассоциации в виде двух последовательно следующих индифферентных стимулов (звук – свет или вертушка – прерывистый шум) было показано, что при отсутствии на них ориентировочных реакций ее формирование идет плохо. В этих же опытах по ОР судили о сформированности ассоциации. Когда собака вырабатывала связь между звуком и светом, можно было видеть, что она поворачивает голову в сторону света еще до его предъявления.
Однако на том этапе изучения ориентировочного рефлекса еще не сложилось полного представления как о его взаимоотношениях с условным рефлексом, так и о физиологических механизмах и структуре самого ориентировочного рефлекса.
П. К. Анохин выделил три формы взаимоотношений ориентировочного рефлекса с текущей деятельностью. К первой он относит «конфликтные отношения», когда ориентировочная активность оказывает тормозный эффект. Эти отношения детально были изучены в лаборатории И.П. Павлова. Вторая форма отражает ассимиляцию ориентировочной реакции оминирующей деятельностью. В этом случае ОР не тормозит, а усиливает текущую деятельность. Например, в ситуации оборонительной доминанты появление нового стимула вызывает не ориентировочный, а оборонительный рефлекс. К третьей форме отношений относится трансформация ОР в поведенческую реакцию, связанную со скрытой, внешне непроявляющейся доминантой. Анализируя эти данные, П.К. Анохин [1968] пришел к выводу том, что эффект ОР существенно зависит от предыдущего опыта животного и от ранее сформировавшихся и зафиксированных в памяти доминант.
Открытие в 1929 г. Г. Бергером альфа-ритма человека и последующее совершенствование техники записи волновой активности мозга позволили связать ОР с депрессией альфа-ритма. Важное значение для изучения ОР имело открытие Г. Моруцци и X. Мэгуном (1949) ретикулярной активирующей системы мозга, позволившее связать блокаду альфа-ритма с реакцией активации, регулируемой этой системой. Наиболее всесторонне ориентировочный рефлекс был исследован в работах Е.Н. Соколова и его коллег.
Полиграфическая регистрация электроэнцефалографических, вегетативных и двигательных реакций животных и человека в условиях многократного повторения индифферентных стимулов привела к представлению об ориентировочном рефлексе как о многокомпонентной реакции активации. Е.Н. Соколов [I960], а затем Р. Линн [Lynn, 1966] описали систему компонентов ОР. На уровне неокор-текса ОР представлен ЭЭГ-реакцией активации (блокадой альфа-ритма, усилением бетаи гаммаколебаний), коррелирующей с увеличением возбудимости, реактивности и лабильности у нейронов коры. К вегетативным компонентам ОР относят расширение зрачков, увеличение кожной проводимости, снижение ЧСС, изменение дыхания, увеличение дыхательной аритмии в частотном спектре ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной сфере ориентировочный рефлекс представлен поворотом головы, движением глаз, настораживанием ушей в направлении нового стимула и увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным реакциям. Под влиянием ОР сенсорная чувствительность анализаторов, измеряемая по субъективным реакциям, увеличивается, что получило название сенсорного компонента ориентировочного рефлекса.
Характер электроэнцефалографических, вегетативных и моторных реакций во время ОР позволяет понять суть рефлекса «Что такое?». Ориентировочный рефлекс обеспечивает лучшее восприятие вызвавшего его стимула и готовность быстро реагировать, если того потребует ситуация. Ради этого прерывается текущая деятельность и ресурсы внимания обращаются на новый стимул.
Отличительной особенностью ОР является его привыкание или угашение, развивающееся при повторении индифферентного стимула. Е.Н. Соколов [1958] выделил две формы ориентировочной реакции: генерализованную и локальную, которые выражают разные этапы процесса угашения ОР. В зависимости от длительности ОР различают тонический и фазический ориентировочный рефлекс.
Первоначально новый стимул вызывает генерализованный ОР, связанный с возбуждением ретикулярной формации ствола мозга. Он характеризуется ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного времени (тоническая форма ОР, часто возникающая параллельно с генерализованным ОР; см. гл. 8). Ее легко увидеть у субъекта, находящегося в дремотном состоянии. Тогда говорят о реакции пробуждения (arousal reaction). Подобную реакцию также наблюдали Шарплесс и Джаспер [Sharpless, Jasper, 1956] у кошек и назвали ее тонической реакцией пробуждения. Генерализованный ориентировочный рефлекс быстро угасает, обычно после 10–15 применений индифферентного стимула.
Это позволяет выявить локальный ориентировочный рефлекс, который более устойчив к угашению и требует 30 и более применений стимула. Его генерацию связывают с активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации сохраняется в сенсорной коре, соответствующей модальности повторяемого раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только в зрительной коре, звук – блокаду тау-ритма, регистрируемого от височной области, тактильное и проприоцептивное раздражение при пассивном подъеме пальца руки вызывает депрессию мю-ритма (роландического ритма) локально в сенсомоторной коре. Длительность реакции сокращается, она становится фазической, появляясь только на включение и выключение стимула (фазическая реакция пробуждения по Шарплису и Джасперу). Продолжая повторять стимул, можно добиться полного угашения ОР.
Ориентировочный рефлекс с полным набором компонентов возникает не у всех испытуемых, что выражает их индивидуальные различия. Изучение компонентного состава ОР на звуковой стимул (1000 Гц, 50 дБ) у 100 испытуемых показало, что блокада альфа-ритма в зрительной коре наблюдается у 85 человек, блокада роландического – у 53, появление КГР – у 95, движение глаз возникало у 42 испытуемых, изменение частоты дыхания – у 63, а изменение мышечного тонуса пальца наблюдалось только у одного испытуемого. Все компоненты ОР (за исключением реакции мышц) были представлены только у 11 испытуемых из 100 (Воронин, Соколов, 1960).
Хотя ориентировочный рефлекс, и относится к безусловным рефлексам, он обнаруживает способность к угашению (так как для него специфическим стимулом является «новизна», а специфической реакцией – набор компонентов ориентировочного рефлекса). Это сближает его с условным рефлексом, который сам по себе является продуктом обучения. Именно поэтому угашение ОР рассматривается как особая форма обучения, получившего название негативного обучения, которое ведет к потере способности у данного стимула вызывать ориентировочные реакции.
Угашение ОР характеризуется высокой избирательностью к физическим характеристикам стимула. Это проявляется в том, что даже небольшое изменение любого из параметров повторяющегося стимула ведет к восстановлению ОР. Стимулы, сильно отличающиеся от угашаемого, вызывают более сильные ориентировочные реакции, а стимулы, близкие ему – более слабые. Величина ОР увеличивается пропорционально количеству одновременно измененных параметров стимула.
Для объяснения этого явления Е.Н. Соколов предположил, что в процессе повторения индифферентного стимула без специального подкрепления в мозге формируется его нейрональная модель (см. также гл. 9). Мозг замечает малейшее отклонение от параметров повторяющегося стимула благодаря сравнению с его моделью, хранящейся в памяти, будь то изменение интенсивности, длительности, цвета, формы, частоты и т.д. В нервной модели запечатлеваются не только элементарные, но и сложные признаки, например комплексы раздражителей – совместное или последовательное применение нескольких раздражителей. Исключение одного из элементов комплекса или изменение их порядка вызывают ориентировочные реакции. Нервная модель стимула многомерна, она фиксирует одновременно все его параметры, что доказывается восстановлением ОР при изменении любого физического свойства повторяющегося стимула даже в случае уменьшения интенсивности или укорочения его длительности.
Новизна как специфический тригер безусловного ОР определяется сигналом рассогласования, генерируемым при несовпадении нервной модели стимула с возбуждением от предъявляемого раздражителя. Схема взаимодействия основных функциональных блоков с нервной моделью, по Е.Н. Соколову, объясняющая угашение и восстановление ОР, включает воспринимающее устройство, исполнительное устройство ОР, блок с моделью стимула, компаратор (устройство, в котором осуществляется операция сравнения стимула с сигналом, поступающим от блока с моделью), блок активирующей системы, определяющий уровень активности исполнительного устройства ориентировочного рефлекса. По мере формирования модели она усиливает свое тормозное влияние на неспецифическую систему мозга. Торможение носит избирательный характер, оно ухудшает восприятие только повторяющегося стимула. В случае несовпадения стимула с моделью в компараторе возникает сигнал рассогласования, который активирует исполнительное устройство ОР.
Следует отметить, что некоторые исследователи увеличивают количество факторов, вызывающих ОР. Класс явлений, названных Д. Берлайном «сличительными признаками», которые стимулируют ориентировочно-исследовательское поведение, включает, помимо новизны, удивление, неопределенность, двусмысленность и сложность. Однако, по мнению Р. Линна [Lynn, 1966], они могут быть интерпретированы как разные формы новизны. Так, удивление связано с появлением чего-то нового в последовательности стимулов, а сложность, необычность и двусмысленность представляют новизну в паттерне стимулов.
Изучение нейронных коррелятов ориентировочного рефлекса выявило существование особого класса нейронов – нейронов новизны, которые были обнаружены в разных структурах мозга. Впервые нейроны новизны были найдены в зрительной покрышке лягушки Дж. Летвиным и др. [1964]. Эти клетки переставали отвечать на повторяющееся движение зрительного объекта, совершаемое в одном и том же направлении. Изменение траектории движения восстанавливало ответ нейрона в виде спайковых разрядов. Нейроны новизны были эфферентными, а их длинные аксоны оканчивались в сетчатке. Эти нейроны сходны с «нейронами внимания», выделенными Д. Хьюбелом и др. (Hubel et al., 1959) в слуховой коре кошки. Нейроны со сходными характеристиками были также найдены в ретикулярной формации ствола мозга кошки (Huttenlocher, 1961; Lindsley et al., 1973). Изучение неспецифического таламуса кролика показало существование особой формы нейронной реакции активации, отличной от простого учащения спайковых разрядов. Новые стимулы вызывали десинхронизацию спайковой активности нейрона, которая состояла из разрушения его пачечной активности, коррелирующей с волнами ЭЭГ, и замены ее на рандоминизированные одиночные спайки, появление которых совпадало с блокадой альфа-подобной активности [Данилова, 1970, 1985]. Реакция десинхронизации нейронов таламуса демонстрирует все свойства ориентировочной реакции.
Вначале формирование нервной модели стимула Е.Н. Соколов связывал с функцией корковых нейронов. Однако после открытия О.С. Виноградовой в поле САЗ гиппокампа двух групп нейронов, реакции которых обнаружили характеристики ОР (В-нейроны, реагирующие возбуждением, и Т-нейроны, отвечающие на новый стимул торможением), он связал нервную модель стимула с нейронами гиппокампа.
Е.Н.Соколов [1995] рассматривает эти две группы нейронов гиппокампа как «нейроны новизны» (В-нейроны) и «нейроны тождества» (Т-нейроны). Их совместное действие позволяет подчеркивать новые раздражители и ослаблять действие привычных стимулов за счет противоположного влияния на активирующую и инактивирующую части модулирующей системы мозга. Формирование нервной модели определяется тем, что множество нейронов-детекторов конвергируют на нейроны поля САЗ гиппокампа (нейроны новизны и тождества). Они создают на каждом из них универсальное рецептивное поле, охватывающее всю рецепторную поверхность животного [Соколов, 1981]. При действии стимула происходит селективное включение детекторов. С повторением раздражителя возбуждение детекторов сохраняется прежним. Однако меняются их синаптические контакты на нейронах гиппокампа. Веса задействованных пластических синапсов на гиппокампальных нейронах уменьшаются пропорционально силе приходящего возбуждения. В результате возбудительный ответ нейрона новизны на повторяющийся стимул подавляется и от него регистрируется лишь фоновая активность. С повторением стимула параллельно меняется и реакция нейронов тождества, с которыми детекторы связаны не возбуждающими, а тормозными пластическими синапсами. Их тормозная реакция на новый стимул постепенно уменьшается, и нейрон сохраняет свою фоновую активность. Е.Н. Соколов предположил, что реципрокные сигналы от нейронов новизны и тождества противоположно воздействуют на активирующую и синхронизирующую системы мозга. Новый раздражитель вызывает реакцию активации, так как возбуждает активирующую и тормозит синхронизирующую (инактивирующую) системы мозга. Привычный стимул перестает возбуждать активирующую неспецифическую систему и при этом через нейроны тождества стимулирует неспецифические тормозные влияния.
Таким образом, нервная модель стимула представлена на нейронах новизны и тождества матрицами потенциированных синапсов, связанных со свойствами раздражителя и отражающими его конфигурацию. Новый стимул активирует новые синапсы нейронов новизны и тождества, еще не подвергнувшихся пластическим перестройкам. Это определяет появление ориентировочного рефлекса за счет возбуждения активирующей и подавления инактивирующей систем мозга. Величина ОР увеличивается пропорционально величине различия (рассогласования) между новым стимулом и сформированной нервной моделью [Соколов, 1995].
Дальнейший прогресс в понимании механизма ОР связан с применением компьютерной техники усреднения при изучении потенциалов мозга, связанных с событиями (ССП; см. гл. 16). Значительный вклад в идентификацию и классификацию различных компонентов ССП и в поиски мозговых аналогов сигналов рассогласования внесли работы Р. Наатанена и его коллег. Им выявлена специфическая мозговая волна, названная негативностью рассогласования (HP). Она получена в опытах, в которых в случайном порядке и с короткими межстимульными интервалами, (не более 10–14 с) следовали стандартный (часто повторяющийся, 80–90%) и девиантные (несколько отклоняющиеся от стандарта и редко предъявляемые, 20–10%) стимулы. При этом внимание испытуемого было направлено на чтение интересной книги. Р. Наатанен для извлечения HP предложил процедуру вычитания ССП на стандартный стимул из ССП на девиантный стимул. Им представлено большое количество экспериментальных доказательств связи HP с операцией сличения действующего стимула с короткоживущим (не более 20 с) следом стандартного стимула в памяти. Чем сильнее они отличаются, тем больше амплитуда и короче латенция HP [Наатанен, 1997].
Исследователями выявлено два компонента HP: модально-специфический и фронтальный. Определяя локусы генерации модально-специфической HP для звуковых стимулов с помощью МЭГ, они нашли их в специфической слуховой коре, где, как предполагает Р. Наатанен, хранятся короткоживущие сенсорные следы. Для разных параметров звуковых стимулов (частоты, интенсивности и пространственного расположения) обнаружены разные HP с несовпадающими локусами их генерации. Предполагают, что сенсорно-специфическая HP трансформируется во фронтальную HP.
Процесс сличения стимула с короткоживущим следом памяти, который представлен в HP, протекает автоматически и не требует сознания. В частности, это подтверждается тем, что HP может возникать и на подпороговые девиации. При этом чем больше амплитуда подпороговой HP, тем ниже субъективный порог различения испытуемого. HP рассматривается как начальная фаза обработки информации о сигнале, за которой может следовать фаза непроизвольного внимания (безусловный ориентировочный рефлекс) к девиантному стимулу. Р. Наатанен связывает его с волной Н2б.
HP как выражение сигнала рассогласования отличается от классического ОР, который, как уже упоминалось, также возникает в результате рассогласования нервной модели стимула и наличного возбуждения от раздражителя. Однако в последнем случае время жизни нервной модели значительно больше, что позволяет при угашении ОР и формировании нервной модели использовать длительные межстимульные интервалы порядка нескольких десятков секунд.
Сравнивая HP с классическим ОР, Р.Наатанен [1997] отмечает, что процесс генерации сенсорно-специфической HP имеет очевидное сходство с процессом рассогласования, постулируемым Е.Н. Соколовым для возникновения ОР, и в то же время между ними имеется существенное различие. HP – первое событие, возникающее в активности мозга, которое направлено на инициацию ОР в ответ на физическое изменение стимула, когда стимулы предъявляются с достаточно короткими межстимульными интервалами, чтобы вызвать HP. Появление ОР в начале стимульной последовательности и в ответ на изменение стимула при длительных межстимульных интервалах имеет другой механизм, отличный от вызова ОР на основе HP.
Другой сигнал рассогласования, который более тесно связан с классическим ОР, был выявлен М. Фербатеном и др. [Verbaten et al., 1986]. Для этого был использован не метод усреднения ССП, а процедура удаления шумового компонента, что позволило исследовать динамику одиночных ССП на стимулы, предъявляемые с большими интервалами. Исследователи нашли, что при угашении ОР (его компонента в виде увеличения кожной проводимости пальцев) параллельно идет процесс уменьшения неспецифической части волны Н1. Новый стимул восстанавливает неспецифический компонент h2 и реакцию повышения кожной проводимости. Выделенная часть компонента Н1 демонстрирует неспецифические свойства ОР и зависимость от долгоживущего следа памяти (десятки секунд).
Таким образом, сигналы рассогласования, участвовавшие в генерации непроизвольного ОР, возникают по крайней мере на двух разных типах следовых процессов: короткоживущей и долгоживущей моделях стимула. Следует отметить, что теория ориентировочного рефлекса Е.Н. Соколова, несмотря на ее большую популярность, все же подвергалась критике. Так, отмечалось, что с ее позиций трудно объяснить появление ОР в случаях когнитивного рассогласования, т.е. когда не подтверждается ожидание относительно последовательности стимулов [Underwood, Moray, 1971]. Например, ориентировочная реакция возникает на цифру «9» в последовательности 1, 2, 3, 4, 5, 6, 7, 9. Другое направление критики связано с утверждением о том, что теория ОР Е.Н. Соколова не объясняет появления ОР на значимые стимулы [Bernstein, 1979; Maltzman, 1979]. Сторонники концепции значимости считают, что решающее значение для появления ОР имеют корковые процессы интерпретации и усиления. С этих позиций теория ОР, возникающего на любое физическое изменение стимула, рассматривается как не оринтированная на реальное поведение и деятельность [Bernstein, 1969].
Однако участники полемики относительно значимости ОР не учитывают, что наряду с безусловным ОР Е.Н. Соколов [1958] вводит понятие условного ориентировочного рефлекса, возникающего на значимый сигнальный стимул. Он показал, что придание раздражителю сигнального значения (обычно с помощью инструкции или речевого подкрепления, побуждающих человека отвечать определенной реакцией на определенный стимул) восстанавливает и усиливает ОР на условный сигнал. Эффект усиления ОР сохраняется до тех пор, пока условный рефлекс не достигнет стадии автоматизации. На рис. 10.1 представлено усиление КГР, измеряемого как падение сопротивления кожи, по Фере, при придании звуковому стимулу сигнального значения. По инструкции испытуемый должен был сжимать руку в кулак при предъявлении звука после предварительного угашения на него безусловного ОР. Выработку условного ориентировочного рефлекса можно увидеть при формировании ассоциации: звук 70 дБ – звук 90 дБ (рис. 10.2). После угашения КГР на индифферентный звук 70 дБ присоединение к нему второго стимула – 90 дБ восстанавливает реакцию, которая усиливается по мере сочетаний стимулов. Постепенно КГР-реакция на комплекс уменьшается и исчезает. Однако изолированное применение стимулов вызывает усиленный ОР, выражающий рассогласование стимула со сформированной в памяти моделью комплекса.
Сравнивая форму ССП на редко предъявляемые девиантные стимулы, когда они были релевантными и испытуемый должен был на них как-то реагировать и когда те же стимулы им игнорировались (испытуемый читал книгу), Р. Наатанен выделил среди компонентов ССП особый потенциал, который коррелировал с произвольным вниманием. Эта волна получила название процессной негативности (ПН). Под влиянием инструкции быть внимательным к особым стимулам, ПН усиливается на все стимулы, но особенно на релевантные. Чем короче межстимульный интервал, тем раньше возникает ПН (в среднем ее ЛП около 150 мс, но она может появляться и через 50 мс).

Рис.10.1 . Усиление КГР – компонента ориентировочного рефлекса на звук, на который испытуемый должен был реагировать движением руки (10 – 24):
9 – инструкция; 1–8– индифферентные звуковые стимулы до инструкции. Видно, что усиление фазических КГР-реакций сопровождается тоническим снижением сопротивления кожи, которое сохраняется до 24-го применения стимула. По оси ординат – сопротивление в килоомах [Соколов, 1958]
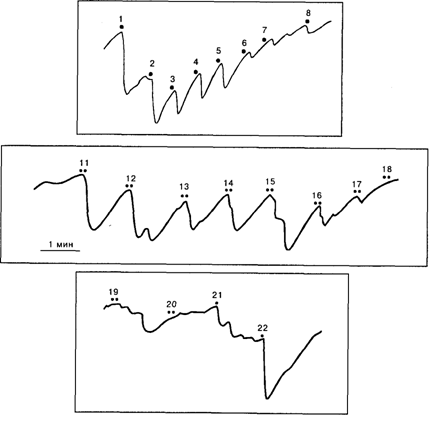
Рис.10.2.Выработка условного ориентировочного рефлекса при формировании ассоциации двух звуковых стимулов:
1-8 — звук 1000 Гц, 70 дБ, 5 с; 11–20 — комплекс: 70 дБ — 90 дБ; 21 — 70 дБ, 22– 90 дБ. По оси ординат – сопротивление в килоомах [Соколов, 1958]
ПН не тождественна Н 1, хотя и совпадает с ней по латенции. Критическим признаком, отличающим Н 1 от ПН, является его инверсия при смещении активного электрода по слуховой коре относительно мостадоидов (сосцевидных отростков). Объясняя ПН, Р. Наатанен [1997] вводит понятие «следа селективного внимания», который связан с памятью. Он полагает, что произвольное внимание актуализирует в сенсорной памяти (по-видимому, во вторичной ассоциативной коре) след релевантного стимула. «След внимания» понимается им как механизм опознания стимула, так как входной сигнал всякий раз сличается со следом релевантного стимула, который непрерывно поддерживает в активном состоянии произвольное внимание. Чем ближе стимул к релевантному, тем больше ПН. Р. Наатанен выделяет раннюю ПН с локусом в ассоциативной коре, выражающей завершение процесса сличения, и позднюю ПН с фронтальной локализацией. Последняя отражает произвольное усилие, направленное на дальнейшую обработку стимула, включающее произвольный ОР на значимый стимул.
Суммируя результаты собственных исследований с данными Р. Наатанена и его коллег, Е.Н. Соколов [1997] выделяет два типа энграмм, с которыми связано возникновение непроизвольного и произвольного ОР. Когда единицы кратковременной памяти подключены к нейронам новизны и тождества прямо в виде потенциированных синапсов от детекторов-признаков, сигнал рассогласования, возникающий при несовпадении стимула с энграммой, определяет генерацию непроизвольного ОР. Его сигнал рассогласования представлен в HP и неспецифическом компоненте Н 1. Если же к нейронам новизны и тождества подключены единицы долговременной памяти и они обеспечивают произвольное удержание шаблона, то совпадение стимула с шаблоном порождает сигнал согласования или совпадения. В терминах нейронных популяций он представлен ПН. Сигнал совпадения достигает нейронов активирующей системы и вызывает условный ОР, направленный на значимый стимул. В случае несовпадения стимула с шаблоном сигнал совпадения и ПН не возникают и ОР на значимый стимул не появляется. При этом стимулы, отличающиеся от релевантного, вызывают безусловный ОР, непроизвольное внимание, которое представлено негативностью рассогласования и неспецифическим компонентом Н1.
Ориентировочный рефлекс
Ориентировочный рефлекс, или рефлекс «Что такое?» был открыт И.П. Павловым. Он описал его как комплекс двигательных реакций, который возникал на неожиданное появление нового стимула. Собака поворачивала голову, глаза, настораживала уши в направлении нового раздражителя. В работах Ю. Конорского [1970] этот комплекс соматических реакций получил название рефлекса прицеливания.
Рассматривая ориентировочный рефлекс как реакцию, направленную на восприятие нового стимула, И.П. Павлов обратил основное внимание на его тормозную, прерывающую функцию в отношении текущей условно-рефлекторной деятельности. Во время ориентировочной реакции условный рефлекс либо полностью подавлялся, либо его выполнение ухудшалось. Это свойство ориентировочной реакции нарушать текущую деятельность, в том числе и процесс обучения (выработку условного рефлекса), привело И.П. Павлова к мысли о необходимости построить исследовательский корпус в виде «Башни молчания». Это позволило бы исключить тормозные эффекты ориентировочного рефлекса (ОР) на формирование и исполнение условных рефлексов.
Хотя вначале ОР и представлялся И. П. Павлову и его коллегам преимущественно как внешний тормоз условного рефлекса, они описали многие его особенности, которые выходили за пределы тормозной функции. Так был открыт растормаживающий эффект ориентировочной реакции. Он состоял в том, что ранее выработанные, а затем угашенные условные связи под влиянием ОР вновь восстанавливались. Сегодня мы интерпретировали бы эти данные как способность восстанавливать подавленные связи в процедурной долговременной памяти. В лаборатории И.П. Павлова также были получены первые факты о влиянии ОР на формирование условного рефлекса. Во время процедуры выработки условной связи, а также при формировании ассоциации между двумя индифферентными раздражителями было отмечено усиление или появление ориентировочных реакций. В опытах И.О. Нарбутович и Н.А. Подкопаева [1936] с выработкой у собаки ассоциации в виде двух последовательно следующих индифферентных стимулов (звук – свет или вертушка – прерывистый шум) было показано, что при отсутствии на них ориентировочных реакций её формирование идёт плохо. В этих же опытах по ОР судили о сформированности ассоциации. Когда собака вырабатывала связь между звуком и светом, можно было видеть, что она поворачивает голову в сторону света ещё до его предъявления.
Однако на том этапе изучения ориентировочного рефлекса ещё не сложилось полного представления как о его взаимоотношениях с условным рефлексом, так и о физиологических механизмах и структуре самого ориентировочного рефлекса.
П. К. Анохин выделил три формы взаимоотношений ориентировочного рефлекса с текущей деятельностью. К первой он относит «конфликтные отношения», когда ориентировочная активность оказывает тормозный эффект. Эти отношения детально были изучены в лаборатории И.П. Павлова. Вторая форма отражает ассимиляцию ориентировочной реакции оминирующей деятельностью. В этом случае ОР не тормозит, а усиливает текущую деятельность. Например, в ситуации оборонительной доминанты появление нового стимула вызывает не ориентировочный, а оборонительный рефлекс. К третьей форме отношений относится трансформация ОР в поведенческую реакцию, связанную со скрытой, внешне непроявляющейся доминантой. Анализируя эти данные, П.К. Анохин [1968] пришёл к выводу том, что эффект ОР существенно зависит от предыдущего опыта животного и от ранее сформировавшихся и зафиксированных в памяти доминант.
Открытие в 1929 г. Г. Бергером альфа-ритма человека и последующее совершенствование техники записи волновой активности мозга позволили связать ОР с депрессией альфа-ритма. Важное значение для изучения ОР имело открытие Г. Моруцци и X. Мэгуном (1949) ретикулярной активирующей системы мозга, позволившее связать блокаду альфа-ритма с реакцией активации, регулируемой этой системой. Наиболее всесторонне ориентировочный рефлекс был исследован в работах Е.Н. Соколова и его коллег.
Полиграфическая регистрация электроэнцефалографических, вегетативных и двигательных реакций животных и человека в условиях многократного повторения индифферентных стимулов привела к представлению об ориентировочном рефлексе как о многокомпонентной реакции активации. Е.Н. Соколов [I960], а затем Р. Линн [Lynn, 1966] описали систему компонентов ОР. На уровне неокор-текса ОР представлен ЭЭГ-реакцией активации (блокадой альфа-ритма, усилением бетаи гаммаколебаний), коррелирующей с увеличением возбудимости, реактивности и лабильности у нейронов коры. К вегетативным компонентам ОР относят расширение зрачков, увеличение кожной проводимости, снижение ЧСС, изменение дыхания, увеличение дыхательной аритмии в частотном спектре ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной сфере ориентировочный рефлекс представлен поворотом головы, движением глаз, настораживанием ушей в направлении нового стимула и увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным реакциям. Под влиянием ОР сенсорная чувствительность анализаторов, измеряемая по субъективным реакциям, увеличивается, что получило название сенсорного компонента ориентировочного рефлекса.
Характер электроэнцефалографических, вегетативных и моторных реакций во время ОР позволяет понять суть рефлекса «Что такое?». Ориентировочный рефлекс обеспечивает лучшее восприятие вызвавшего его стимула и готовность быстро реагировать, если того потребует ситуация. Ради этого прерывается текущая деятельность и ресурсы внимания обращаются на новый стимул.
Отличительной особенностью ОР является его привыкание или угашение, развивающееся при повторении индифферентного стимула. Е.Н. Соколов выделил две формы ориентировочной реакции: генерализованную и локальную, которые выражают разные этапы процесса угашения ОР. В зависимости от длительности ОР различают тонический и фазический ориентировочный рефлекс.
Первоначально новый стимул вызывает генерализованный ОР, связанный с возбуждением ретикулярной формации ствола мозга. Он характеризуется ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного времени (тоническая форма ОР, часто возникающая параллельно с генерализованным ОР; см. гл. 8). Её легко увидеть у субъекта, находящегося в дремотном состоянии. Тогда говорят о реакции пробуждения (arousal reaction). Подобную реакцию также наблюдали Шарплесс и Джаспер [Sharpless, Jasper, 1956] у кошек и назвали её тонической реакцией пробуждения. Генерализованный ориентировочный рефлекс быстро угасает, обычно после 10–15 применений индифферентного стимула.
Это позволяет выявить локальный ориентировочный рефлекс, который более устойчив к угашению и требует 30 и более применений стимула. Его генерацию связывают с активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации сохраняется в сенсорной коре, соответствующей модальности повторяемого раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только в зрительной коре, звук – блокаду тау-ритма, регистрируемого от височной области, тактильное и проприоцептивное раздражение при пассивном подъёме пальца руки вызывает депрессию мю-ритма (роландического ритма) локально в сенсомоторной коре. Длительность реакции сокращается, она становится фазической, появляясь только на включение и выключение стимула (фазическая реакция пробуждения по Шарплису и Джасперу). Продолжая повторять стимул, можно добиться полного угашения ОР.
Ориентировочный рефлекс с полным набором компонентов возникает не у всех испытуемых, что выражает их индивидуальные различия. Изучение компонентного состава ОР на звуковой стимул (1000 Гц, 50 дБ) у 100 испытуемых показало, что блокада альфа-ритма в зрительной коре наблюдается у 85 человек, блокада роландического – у 53, появление КГР – у 95, движение глаз возникало у 42 испытуемых, изменение частоты дыхания – у 63, а изменение мышечного тонуса пальца наблюдалось только у одного испытуемого. Все компоненты ОР (за исключением реакции мышц) были представлены только у 11 испытуемых из 100.
Хотя ориентировочный рефлекс, и относится к безусловным рефлексам, он обнаруживает способность к угашению (так как для него специфическим стимулом является «новизна», а специфической реакцией – набор компонентов ориентировочного рефлекса). Это сближает его с условным рефлексом, который сам по себе является продуктом обучения. Именно поэтому угашение ОР рассматривается как особая форма обучения, получившего название негативного обучения, которое ведёт к потере способности у данного стимула вызывать ориентировочные реакции.
Угашение ОР характеризуется высокой избирательностью к физическим характеристикам стимула. Это проявляется в том, что даже небольшое изменение любого из параметров повторяющегося стимула ведёт к восстановлению ОР. Стимулы, сильно отличающиеся от угашаемого, вызывают более сильные ориентировочные реакции, а стимулы, близкие ему – более слабые. Величина ОР увеличивается пропорционально количеству одновременно изменённых параметров стимула.
Для объяснения этого явления Е.Н. Соколов предположил, что в процессе повторения индифферентного стимула без специального подкрепления в мозге формируется его нейрональная модель (см. также гл. 9). Мозг замечает малейшее отклонение от параметров повторяющегося стимула благодаря сравнению с его моделью, хранящейся в памяти, будь то изменение интенсивности, длительности, цвета, формы, частоты и т.д. В нервной модели запечатлеваются не только элементарные, но и сложные признаки, например комплексы раздражителей – совместное или последовательное применение нескольких раздражителей. Исключение одного из элементов комплекса или изменение их порядка вызывают ориентировочные реакции. Нервная модель стимула многомерна, она фиксирует одновременно все его параметры, что доказывается восстановлением ОР при изменении любого физического свойства повторяющегося стимула даже в случае уменьшения интенсивности или укорочения его длительности.
Новизна как специфический тригер безусловного ОР определяется сигналом рассогласования, генерируемым при несовпадении нервной модели стимула с возбуждением от предъявляемого раздражителя. Схема взаимодействия основных функциональных блоков с нервной моделью, по Е.Н. Соколову, объясняющая угашение и восстановление ОР, включает воспринимающее устройство, исполнительное устройство ОР, блок с моделью стимула, компаратор (устройство, в котором осуществляется операция сравнения стимула с сигналом, поступающим от блока с моделью), блок активирующей системы, определяющий уровень активности исполнительного устройства ориентировочного рефлекса. По мере формирования модели она усиливает своё тормозное влияние на неспецифическую систему мозга. Торможение носит избирательный характер, оно ухудшает восприятие только повторяющегося стимула. В случае несовпадения стимула с моделью в компараторе возникает сигнал рассогласования, который активирует исполнительное устройство ОР.
Следует отметить, что некоторые исследователи увеличивают количество факторов, вызывающих ОР. Класс явлений, названных Д. Берлайном «сличительными признаками», которые стимулируют ориентировочно-исследовательское поведение, включает, помимо новизны, удивление, неопределённость, двусмысленность и сложность. Однако, по мнению Р. Линна [Lynn, 1966], они могут быть интерпретированы как разные формы новизны. Так, удивление связано с появлением чего-то нового в последовательности стимулов, а сложность, необычность и двусмысленность представляют новизну в паттерне стимулов.
Изучение нейронных коррелятов ориентировочного рефлекса выявило существование особого класса нейронов – нейронов новизны, которые были обнаружены в разных структурах мозга. Впервые нейроны новизны были найдены в зрительной покрышке лягушки Дж. Летвиным и др. [1964]. Эти клетки переставали отвечать на повторяющееся движение зрительного объекта, совершаемое в одном и том же направлении. Изменение траектории движения восстанавливало ответ нейрона в виде спайковых разрядов. Нейроны новизны были эфферентными, а их длинные аксоны оканчивались в сетчатке. Эти нейроны сходны с «нейронами внимания», выделенными Д. Хьюбелом и др. (Hubel et al., 1959) в слуховой коре кошки. Нейроны со сходными характеристиками были также найдены в ретикулярной формации ствола мозга кошки (Huttenlocher, 1961; Lindsley et al., 1973). Изучение неспецифического таламуса кролика показало существование особой формы нейронной реакции активации, отличной от простого учащения спайковых разрядов. Новые стимулы вызывали десинхронизацию спайковой активности нейрона, которая состояла из разрушения его пачечной активности, коррелирующей с волнами ЭЭГ, и замены её на рандоминизированные одиночные спайки, появление которых совпадало с блокадой альфа-подобной активности [Данилова, 1970, 1985]. Реакция десинхронизации нейронов таламуса демонстрирует все свойства ориентировочной реакции.
Вначале формирование нервной модели стимула Е.Н. Соколов связывал с функцией корковых нейронов. Однако после открытия О.С. Виноградовой в поле САЗ гиппокампа двух групп нейронов, реакции которых обнаружили характеристики ОР (В-нейроны, реагирующие возбуждением, и Т-нейроны, отвечающие на новый стимул торможением), он связал нервную модель стимула с нейронами гиппокампа.
Е.Н.Соколов [1995] рассматривает эти две группы нейронов гиппокампа как «нейроны новизны» (В-нейроны) и «нейроны тождества» (Т-нейроны). Их совместное действие позволяет подчёркивать новые раздражители и ослаблять действие привычных стимулов за счёт противоположного влияния на активирующую и инактивирующую части модулирующей системы мозга. Формирование нервной модели определяется тем, что множество нейронов-детекторов конвергируют на нейроны поля САЗ гиппокампа (нейроны новизны и тождества). Они создают на каждом из них универсальное рецептивное поле, охватывающее всю рецепторную поверхность животного [Соколов, 1981]. При действии стимула происходит селективное включение детекторов. С повторением раздражителя возбуждение детекторов сохраняется прежним. Однако меняются их синаптические контакты на нейронах гиппокампа. Веса задействованных пластических синапсов на гиппокампальных нейронах уменьшаются пропорционально силе приходящего возбуждения. В результате возбудительный ответ нейрона новизны на повторяющийся стимул подавляется и от него регистрируется лишь фоновая активность. С повторением стимула параллельно меняется и реакция нейронов тождества, с которыми детекторы связаны не возбуждающими, а тормозными пластическими синапсами. Их тормозная реакция на новый стимул постепенно уменьшается, и нейрон сохраняет свою фоновую активность. Е.Н. Соколов предположил, что реципрокные сигналы от нейронов новизны и тождества противоположно воздействуют на активирующую и синхронизирующую системы мозга. Новый раздражитель вызывает реакцию активации, так как возбуждает активирующую и тормозит синхронизирующую (инактивирующую) системы мозга. Привычный стимул перестаёт возбуждать активирующую неспецифическую систему и при этом через нейроны тождества стимулирует неспецифические тормозные влияния.
Таким образом, нервная модель стимула представлена на нейронах новизны и тождества матрицами потенциированных синапсов, связанных со свойствами раздражителя и отражающими его конфигурацию. Новый стимул активирует новые синапсы нейронов новизны и тождества, ещё не подвергнувшихся пластическим перестройкам. Это определяет появление ориентировочного рефлекса за счёт возбуждения активирующей и подавления инактивирующей систем мозга. Величина ОР увеличивается пропорционально величине различия (рассогласования) между новым стимулом и сформированной нервной моделью [Соколов, 1995].
Дальнейший прогресс в понимании механизма ОР связан с применением компьютерной техники усреднения при изучении потенциалов мозга, связанных с событиями (ССП; см. гл. 16). Значительный вклад в идентификацию и классификацию различных компонентов ССП и в поиски мозговых аналогов сигналов рассогласования внесли работы Р. Наатанена и его коллег. Им выявлена специфическая мозговая волна, названная негативностью рассогласования (HP). Она получена в опытах, в которых в случайном порядке и с короткими межстимульными интервалами, (не более 10–14 с) следовали стандартный (часто повторяющийся, 80–90%) и девиантные (несколько отклоняющиеся от стандарта и редко предъявляемые, 20–10%) стимулы. При этом внимание испытуемого было направлено на чтение интересной книги. Р. Наатанен для извлечения HP предложил процедуру вычитания ССП на стандартный стимул из ССП на девиантный стимул. Им представлено большое количество экспериментальных доказательств связи HP с операцией сличения действующего стимула с короткоживущим (не более 20 с) следом стандартного стимула в памяти. Чем сильнее они отличаются, тем больше амплитуда и короче латенция HP [Наатанен, 1997].
Исследователями выявлено два компонента HP: модально-специфический и фронтальный. Определяя локусы генерации модально-специфической HP для звуковых стимулов с помощью МЭГ, они нашли их в специфической слуховой коре, где, как предполагает Р. Наатанен, хранятся короткоживущие сенсорные следы. Для разных параметров звуковых стимулов (частоты, интенсивности и пространственного расположения) обнаружены разные HP с несовпадающими локусами их генерации. Предполагают, что сенсорно-специфическая HP трансформируется во фронтальную HP.
Процесс сличения стимула с короткоживущим следом памяти, который представлен в HP, протекает автоматически и не требует сознания. В частности, это подтверждается тем, что HP может возникать и на подпороговые девиации. При этом чем больше амплитуда подпороговой HP, тем ниже субъективный порог различения испытуемого. HP рассматривается как начальная фаза обработки информации о сигнале, за которой может следовать фаза непроизвольного внимания (безусловный ориентировочный рефлекс) к девиантному стимулу. Р. Наатанен связывает его с волной Н.
HP как выражение сигнала рассогласования отличается от классического ОР, который, как уже упоминалось, также возникает в результате рассогласования нервной модели стимула и наличного возбуждения от раздражителя. Однако в последнем случае время жизни нервной модели значительно больше, что позволяет при угашении ОР и формировании нервной модели использовать длительные межстимульные интервалы порядка нескольких десятков секунд.
Сравнивая HP с классическим ОР, Р.Наатанен [1997] отмечает, что процесс генерации сенсорно-специфической HP имеет очевидное сходство с процессом рассогласования, постулируемым Е.Н. Соколовым для возникновения ОР, и в то же время между ними имеется существенное различие. HP – первое событие, возникающее в активности мозга, которое направлено на инициацию ОР в ответ на физическое изменение стимула, когда стимулы предъявляются с достаточно короткими межстимульными интервалами, чтобы вызвать HP. Появление ОР в начале стимульной последовательности и в ответ на изменение стимула при длительных межстимульных интервалах имеет другой механизм, отличный от вызова ОР на основе HP.
Другой сигнал рассогласования, который более тесно связан с классическим ОР, был выявлен М. Фербатеном и др. [Verbaten et al., 1986]. Для этого был использован не метод усреднения ССП, а процедура удаления шумового компонента, что позволило исследовать динамику одиночных ССП на стимулы, предъявляемые с большими интервалами. Исследователи нашли, что при угашении ОР (его компонента в виде увеличения кожной проводимости пальцев) параллельно идёт процесс уменьшения неспецифической части волны Н1. Новый стимул восстанавливает неспецифический компонент h2 и реакцию повышения кожной проводимости. Выделенная часть компонента Н1 демонстрирует неспецифические свойства ОР и зависимость от долгоживущего следа памяти (десятки секунд).
Таким образом, сигналы рассогласования, участвовавшие в генерации непроизвольного ОР, возникают по крайней мере на двух разных типах следовых процессов: короткоживущей и долгоживущей моделях стимула. Следует отметить, что теория ориентировочного рефлекса Е.Н. Соколова, несмотря на её большую популярность, все же подвергалась критике. Так, отмечалось, что с её позиций трудно объяснить появление ОР в случаях когнитивного рассогласования, т.е. когда не подтверждается ожидание относительно последовательности стимулов [Underwood, Moray, 1971]. Например, ориентировочная реакция возникает на цифру «9» в последовательности 1, 2, 3, 4, 5, 6, 7, 9. Другое направление критики связано с утверждением о том, что теория ОР Е.Н. Соколова не объясняет появления ОР на значимые стимулы [Bernstein, 1979; Maltzman, 1979]. Сторонники концепции значимости считают, что решающее значение для появления ОР имеют корковые процессы интерпретации и усиления. С этих позиций теория ОР, возникающего на любое физическое изменение стимула, рассматривается как не оринтированная на реальное поведение и деятельность [Bernstein, 1969].
Однако участники полемики относительно значимости ОР не учитывают, что наряду с безусловным ОР Е.Н. Соколов [1958] вводит понятие условного ориентировочного рефлекса, возникающего на значимый сигнальный стимул. Он показал, что придание раздражителю сигнального значения (обычно с помощью инструкции или речевого подкрепления, побуждающих человека отвечать определённой реакцией на определённый стимул) восстанавливает и усиливает ОР на условный сигнал. Эффект усиления ОР сохраняется до тех пор, пока условный рефлекс не достигнет стадии автоматизации. На рис. 10.1 представлено усиление КГР, измеряемого как падение сопротивления кожи, по Фере, при придании звуковому стимулу сигнального значения. По инструкции испытуемый должен был сжимать руку в кулак при предъявлении звука после предварительного угашения на него безусловного ОР. Выработку условного ориентировочного рефлекса можно увидеть при формировании ассоциации: звук 70 дБ – звук 90 дБ (рис. 10.2). После угашения КГР на индифферентный звук 70 дБ присоединение к нему второго стимула – 90 дБ восстанавливает реакцию, которая усиливается по мере сочетаний стимулов. Постепенно КГР-реакция на комплекс уменьшается и исчезает. Однако изолированное применение стимулов вызывает усиленный ОР, выражающий рассогласование стимула со сформированной в памяти моделью комплекса.
Сравнивая форму ССП на редко предъявляемые девиантные стимулы, когда они были релевантными и испытуемый должен был на них как-то реагировать и когда те же стимулы им игнорировались (испытуемый читал книгу), Р. Наатанен выделил среди компонентов ССП особый потенциал, который коррелировал с произвольным вниманием. Эта волна получила название процессной негативности (ПН). Под влиянием инструкции быть внимательным к особым стимулам, ПН усиливается на все стимулы, но особенно на релевантные. Чем короче межстимульный интервал, тем раньше возникает ПН (в среднем её ЛП около 150 мс, но она может появляться и через 50 мс).
ПН не тождественна Н 1, хотя и совпадает с ней по латенции. Критическим признаком, отличающим Н 1 от ПН, является его инверсия при смещении активного электрода по слуховой коре относительно мостадоидов (сосцевидных отростков). Объясняя ПН, Р. Наатанен [1997] вводит понятие «следа селективного внимания», который связан с памятью. Он полагает, что произвольное внимание актуализирует в сенсорной памяти (по-видимому, во вторичной ассоциативной коре) след релевантного стимула. «След внимания» понимается им как механизм опознания стимула, так как входной сигнал всякий раз сличается со следом релевантного стимула, который непрерывно поддерживает в активном состоянии произвольное внимание. Чем ближе стимул к релевантному, тем больше ПН. Р. Наатанен выделяет раннюю ПН с локусом в ассоциативной коре, выражающей завершение процесса сличения, и позднюю ПН с фронтальной локализацией. Последняя отражает произвольное усилие, направленное на дальнейшую обработку стимула, включающее произвольный ОР на значимый стимул.
Суммируя результаты собственных исследований с данными Р. Наатанена и его коллег, Е.Н. Соколов [1997] выделяет два типа энграмм, с которыми связано возникновение непроизвольного и произвольного ОР. Когда единицы кратковременной памяти подключены к нейронам новизны и тождества прямо в виде потенциированных синапсов от детекторов-признаков, сигнал рассогласования, возникающий при несовпадении стимула с энграммой, определяет генерацию непроизвольного ОР. Его сигнал рассогласования представлен в HP и неспецифическом компоненте Н 1. Если же к нейронам новизны и тождества подключены единицы долговременной памяти и они обеспечивают произвольное удержание шаблона, то совпадение стимула с шаблоном порождает сигнал согласования или совпадения. В терминах нейронных популяций он представлен ПН. Сигнал совпадения достигает нейронов активирующей системы и вызывает условный ОР, направленный на значимый стимул. В случае несовпадения стимула с шаблоном сигнал совпадения и ПН не возникают и ОР на значимый стимул не появляется. При этом стимулы, отличающиеся от релевантного, вызывают безусловный ОР, непроизвольное внимание, которое представлено негативностью рассогласования и неспецифическим компонентом Н1.
Ориентировочный рефлекс. Общие закономерности формирования врожденных форм поведения
Особое место среди безусловных рефлексов занимает ориентировочный рефлекс. Это рефлекс на новизну. Он возникает в ответ на любое достаточно быстро происходящее изменение окружающей среды  . Ориентировочным рефлексом называют безусловно-рефлекторные непроизвольные реакции, сопровождающиеся резким повышением внимания и тонуса мышц и вызванные неожиданным или новым для организма раздражителем. Ориентировочный рефлекс выражается внешне в настораживании, прислушивании, обнюхивании, повороте глаз и головы, а иногда и всего тела в сторону появившегося нового раздражителя. Осуществление этого рефлекса обеспечивает лучшее восприятие действующего агента и имеет важное приспособительное значение.
. Ориентировочным рефлексом называют безусловно-рефлекторные непроизвольные реакции, сопровождающиеся резким повышением внимания и тонуса мышц и вызванные неожиданным или новым для организма раздражителем. Ориентировочный рефлекс выражается внешне в настораживании, прислушивании, обнюхивании, повороте глаз и головы, а иногда и всего тела в сторону появившегося нового раздражителя. Осуществление этого рефлекса обеспечивает лучшее восприятие действующего агента и имеет важное приспособительное значение.
Ориентировочный рефлекс часто называют рефлексом настораживания, тревоги, удивления, а И.П. Павлов назвал его рефлексом «Что такое?». Чаще всего ориентировочный рефлекс возникает при неожиданном раздражении зрительных, слуховых или обонятельных рецепторов. Реакция эта врожденная и не исчезает при полном удалении коры больших полушарий у животных. Ее наблюдают у детей с недоразвитыми полушариями – анэнцефалов. Отличием ориентировочного рефлекса от других безусловно-рефлекторных реакций является то, что он сравнительно быстро ослабляется и угасает при повторных применениях одного и того же раздражителя. Эта особенность ориентировочного рефлекса зависит от коры больших полушарий.
Ориентировочный рефлекс проявляется целым комплексом реакций. В развитии ориентировочного рефлекса выделяют три фазы. Первая фаза выражается в прекращении текущей деятельности с фиксацией позы. Возникает общее торможение («стоп-реакция») в ответ на появление любого достаточно сильного постороннего раздражителя с неизвестным сигнальным значением.
Вторая фаза начинается, когда состояние «стоп-реакции» переходит в реакцию вздрагивания, или общей активности. В эту фазу весь организм приводится в состояние рефлекторной готовности к возможной встрече с чрезвычайной ситуацией. Мобилизация самозащиты организма при встрече с новым или неожиданным раздражителем выражается в общем повышении тонуса всей скелетной мускулатуры, повороте головы и глаз в направлении раздражителя.
Третья фаза ориентировочного рефлекса начинается с фиксации раздражителя органами чувств. Затем происходит развертывание процесса анализа поступающей от рецепторов информации о раздражителе и принятие решения об ответной реакции организма.
Ориентировочный рефлекс входит в структуру ориентировочного поведения, или ориентировочно-исследовательской деятельности. Исследовательская активность в новой обстановке может быть направлена как на освоение новизны, удовлетворение любопытства, оценку ситуации, так и на поиск раздражителя, способного удовлетворить имеющуюся потребность.
Ориентировочный рефлекс помогает определить значимость раздражителя, в связи с чем повышается чувствительность анализаторов для наилучшего восприятия раздражителя. Таким образом, любой вид анализа раздражителей происходит при активном участии ориентировочного рефлекса.
Осуществление ориентировочного рефлекса является результатом динамического взаимодействия различных структур коры больших полушарий и подкорковых образований головного мозга, что находит отражение в изменении ритма электроэнцефалограммы. Ориентировочная реакция возникает в результате сличения (сопоставления) нового или неожиданного раздражителя со следом («нервной моделью»), оставленным в нервной системе предшествующими раздражителями. Если раздражитель и след совпадают, то ориентировочный рефлекс не возникает. Если же поступающая от органов чувств информация о раздражителе не совпадает с хранящимся в памяти следом («нервной моделью» раздражителя), то ориентировочная реакция возникает и оказывается тем интенсивнее, чем более выражено такое рассогласование.
Формирование врожденных форм поведения характеризуется следующими общими закономерностями.
1) Каждый компонент безусловно-рефлекторной деятельности развертывается на основе внутренней потребности, вызвавшей доминирующую мотивацию, и действия специальных ключевых факторов внешней среды.
2) Генетически обусловленные компоненты поведения характеризуются жестким программированием этапных и конечных результатов поведения, удовлетворяющих главные потребности организма.
3) Обязательным условием удовлетворения ведущей потребности организма при инстинктивной деятельности является достижение всех этапных результатов и сравнение их с генетически запрограммированными. Только после получения полноценной информации о предыдущем этапном результате животные осуществляют деятельность, направленную на достижение следующего результата.
4) Для формирования инстинктивной деятельности необходимы стабильные на протяжении многих поколений условия существования животных организмов.
5) Инстинктивное поведение мало использует механизмы индивидуального обучения. Как правило, инстинктивное поведение более выражено у животных, вырастающих изолированно от родителей.
Ориентировочный рефлекс, как основа внимания. — Студопедия
Оглавление.
| Введение
1.1.Понятие внимания. Физиологические основы внимания 1.2. Физиологические основы внимания 1.3. Ориентировочный рефлекс, как основа внимания 1.4. Виды внимания 1.5. Факторы, определяющие внимание 1.6. Развитие внимания в онтогенезе человека 1.7. Внимание в учебном процессе 1.8. Методы исследования внимания 1.9. Исследование объема внимания 1.10. Исследование устойчивости внимания 1.11. Исследование распределения внимания 2. Описание диагностических методов исследования внимания Список литературы | 3
4 5 8 11 15 20 27 31 31 32 33 35 41 |
Введение.
Внимание – важное и необходимое условие эффективности всех видов деятельности человека, прежде всего трудовой и учебной. Чем сложнее и ответственнее труд, тем больше требований предъявляет он к вниманию. Внимательность необходима человеку в его повседневной жизни, в быту, в общении с другими людьми, в спорте. Внимание учащихся является одним из основных условий успешной организации учебно-воспитательного процесса. Еще великий русский педагог К.Д. Ушинский, подчеркивая роль внимания в обучении, писал: «…внимание есть именно та дверь, через которую проходит все, что только входит в душу человека из внешнего мира». Приучить детей держать эти двери открытыми есть дело первой важности, на успехе которого основывается успех всего учения. Важнейшей особенностью протекания познавательных процессов является их избирательный, направленный характер. Из множества воздействий окружающего мира человек всегда воспринимает что-то, что-то представляет, размышляет, думает о чем-то. Эту особенность сознания связывают с таким его свойством, как внимание. В отличие от познавательных процессов внимание своего особого содержания не имеет, оно – динамическая сторона всех познавательных процессов. Внимание активизирует нужные и тормозит ненужные в данный момент психологические и физиологические процессы, способствует целенаправленному, организованному отбору поступающей информации, обеспечивает длительную сосредоточенность активности на одном и том же объекте.
Цель работы: изучить основные характеристики внимания и определить его роль в развитии познавательных процессов.
1.1.Понятие внимания. Физиологические основы внимания.
До человека доходит огромное число раздражителей, однако он отбирает самые важные из них и игнорирует остальные. Он потенциально может сделать большое число возможных движений, но выделяет немногие целесообразные движения, входящие в состав его действий, и тормозит остальные. У него возникает большое число ассоциаций, однако он сохраняет лишь немногие, существенные для его деятельности, и абстрагируется от других, мешающих целенаправленному протеканию его мышления
Осуществление отбора нужной информации, обеспечение избирательных программ действий и сохранение постоянного контроля над их протеканием и принято называть вниманием.
Избирательный характер сознательной деятельности, который является функцией внимания, в равной мере проявляется и в нашем восприятии, и в двигательных процессах, и в мышлении.
Если бы такой избирательности не было, количество неотобранной информации было бы так неорганизованно и велико, что никакая деятельность не была бы возможна. Если бы не было торможения всех бесконтрольно всплывающих ассоциаций, организованное мышление, направленное на решение поставленных перед человеком задач, было бы недоступно.
Во всех видах сознательной деятельности должны иметь место:
1) процесс выбора основных, доминирующих процессов, составляющих предмет, на который человек обращает внимание;
2) наличие «фона», состоящего из тех процессов, доступ которых в сознание задержан, но они в любой момент, если появляется соответствующая задача, могут перейти в центр его внимания и стать доминирующими.
Именно в связи с этим принято различать объем внимания, его устойчивость и его колебания.
Под объемом внимания принято понимать то число поступающих сигналов или протекающих ассоциаций, которые могут сохраняться в центре ясного сознания, приобретая доминирующий характер.
Под устойчивостью внимания принято понимать ту длительность, с которой эти выделенные вниманием процессы могут сохранять свой доминирующий характер.
Под колебаниями внимания принято понимать тот циклический характер процесса, при котором определенные содержания сознательной деятельности то приобретают доминирующий характер, то теряют его.[3]
1.2. Физиологические основы внимания.
В течение длительного времени психологи, и физиологи пытались описать механизмы, которые определяют избирательное протекание процессов возбуждения и лежат в основе внимания. Однако эти попытки долго ограничивались лишь указанием на тот или иной фактор и скорее носили описательный характер, чем характер подлинного выделения физиологических механизмов внимания.
Некоторые психологи считали, что направление и объем внимания целиком определяются законами структурного восприятия, поэтому излишне выделять исследование внимания в специальную главу психологии и знания таких законов, как законы «четкости», «структурности» восприятия, достаточно, чтобы исчерпывающе судить о протекании внимания. Такую позицию занимали представители гештальт- психологии, один из которых посвятил этому даже специальную статью, пытаясь доказать тезис о том, что внимания как особой категории психических процессов, отдельных от восприятия, не существует.
Другая группа психологов придерживалась позиций «эмоциональной» теории внимания. Эти психологи полагали, что направление внимания целиком определяется влечениями, потребностями и эмоциями, исчерпываясь законами, и что внимание не должно выделяться в особую категорию психических процессов. Многие американские психологи- бихевиористы практически занимают эту позицию. Наконец, третья группа психологов, подходящих к проблеме с позиции «моторной теории внимания», видит во внимании проявление тех моторных установок, которые лежат в основе каждого волевого акта, и считают, что механизмом внимания являются сигналы мышечных усилий, которыми характеризуется всякое напряжение, вызванное определенной, направленной на известную цель деятельностью.
Легко видеть, что каждая из этих теорий выделяет определенный компонент, входящий в состав внимания, но фактически даже не пытается подойти к вопросу об общих физиологических механизмах, лежащих в основе внимания.
Значительные трудности возникали перед физиологами, которые выдвигали гипотезы об общих физиологических основах внимания.
В течение длительного времени эти попытки носили слишком общий характер и заключались скорее в описании общих условий избирательного протекания возбуждения, чем выделении специальных физиологических механизмов внимания.[3]
Одной из наиболее ранних попыток была гипотеза известного английского физиолога Ч. Шеррингтона, которая в дальнейшем получила широко известное название «теории общего двигательного поля», или «воронки Шеррингтона». Наблюдая факт, что сенсорных нейронов в задних рогах спинного мозга значительно больше, чем двигательных нейронов, Ч. Шеррингтон высказал положение, что не каждый двигательный импульс может дойти до своего двигательного конца, и большое число сенсорных возбуждении имеют свое «общее двигательное поле», что отношение сенсорных и моторных процессов можно уподобить воронке, в широкое отверстие которой входят сенсорные импульсы, а из узкого отверстия выходят двигательные иннервации. Легко видеть, что среди сенсорных импульсов возникает « борьба за общее двигательное поле», в которой побеждают наиболее сильные, наиболее подготовленные или входящие в известную биологическую систему импульсы. Несмотря на то, что Ч. Шеррингтон был одним из первых физиологов, исследовавшим интегративную деятельность мозга и сформулировавшим положение о системном строении физиологических процессов, теория «борьбы за общее двигательное поле» лишь в самых общих чертах приближается к указанию на физиологические механизмы, лежащие в основе внимания.
Такой же общий и скорее метафорический характер имеют и ранние высказывания И. П. Павлова, который уподоблял внимание (и ясное сознание) очагу оптимального возбуждения, движущегося по коре головного мозга наподобие «перемещающегося светового пятна». Идея очага оптимального возбуждения как основы внимания оказалась в дальнейшем очень важной и подводила к некоторым существенным физиологическим механизмам внимания, но, конечно, была слишком общей, чтобы удовлетворительно объяснять эти процессы.
Значительный вклад в анализ физиологических механизмов внимания внес замечательный русский физиолог Л. А. Ухтомский. Согласно его представлениям, возбуждение распределяется по нервной системе неравномерно, и каждая инстинктивная деятельность (как и условно-рефлекторные процессы) может создавать в нервной системе очаги оптимального возбуждения, которые приобретают доминирующий характер. Эти очаги, которые А. А. Ухтомский обозначил термином доминанты, не только господствуют над другими и тормозят существующие наряду с ними другие очаги, но приобретают даже способность усиливаться под влиянием действия посторонних возбуждений.
Так, лягушка, у которой в определенный период создается доминанта обхватывающего рефлекса передних лапок, реагирует на раздражение задних лапок усилением доминирующих обхватывающих движений передних лапок. Эта способность доминанты тормозить побочные рефлексы и даже усиливаться под влиянием посторонних раздражителей была оценена А. А. Ухтомским как процесс, напоминающий внимание, и именно это дало ему основание расценивать доминанту как физиологический механизм внимания.[3]
Ориентировочный рефлекс, как основа внимания.
Активирующая ретикулярная система с ее восходящими и нисходящими волокнами является нейрофизиологическим аппаратом, обеспечивающим одну из наиболее важных форм рефлекторной деятельности, известную под названием ориентировочного (или ориентировочно-исследовательского) рефлекса. Его значение для понимания физиологических основ внимания настолько велико, что на нем следует остановиться специально.
Каждый безусловный рефлекс, имеющий в своей основе какое-либо биологически важное для животного воздействие (пищевое, болевое, половое), вызывает избирательную систему ответов на эти раздражители с одновременным торможением всех реакций на побочные. Такой же избирательный характер носят и условные рефлексы. При них одна система реакций, подкрепляемая безусловным раздражителем, доминирует, в то время как все остальные побочные реакции тормозятся. Можно сказать, что как безусловные, так и сформированные на их основе условные рефлексы создают известный доминирующий очаг возбуждения, протекание которого подчиняется доминанте.
Среди всех видов рефлекторной деятельности нужно, однако, выделить один, при котором поведение животного не возбуждается ни одним из перечисленных выше мотивов поведения и который не является ни пищевым, ни оборонительным, ни половым рефлексом. Основой этой деятельности является активная реакция животного на каждое изменение обстановки, которое и вызывает у животного общее оживление и ряд избирательных реакций, направленных на ознакомление с этими изменениями в ситуации. И. П. Павлов назвал этот вид рефлексов «ориентировочными рефлексами», или «рефлексами «что такое?»».
Ориентировочный рефлекс выражается в ряде отчетливых электрофизиологических, сосудистых и двигательных реакций, появляющихся каждый раз, когда в обстановке, окружающей животное, возникает что-нибудь необычное или существенное. К этим реакциям относятся :
• поворот глаз и головы в сторону нового объекта;
• реакция настораживания или прислушивания.
А у человека — появление кожно-гальванической реакции (изменение сопротивляемости кожи к электрическому току или появление собственных электрических потенциалов кожи), сосудистые реакции (сужение сосудов руки с расширением сосудов головы), изменение дыхания, наконец, возникновение явлений «десин-хронизации» в биоэлектрических реакциях мозга, выражающихся в депрессии «альфа ритма» (электрических колебаний 10-12 в сек, характерных для работы мозговой коры в спокойном состоянии). Все эти явления можно наблюдать каждый раз, когда возникает реакция настораживания, или ориентировочный рефлекс, вызываемый появлением нового или существенного для субъекта раздражителя.
Среди ученых нет еще единодушного ответа на вопрос, является ли ориентировочный рефлекс безусловной или условной реакцией.
По своему врожденному характеру ориентировочный рефлекс можно отнести к числу безусловных. Животное отвечает реакцией настораживания на любые новые пли существенные раздражители без всякого обучения; но этому признаку ориентировочный рефлекс относится к числу безусловных, врожденных реакции организма. Наличие особых нейронов, которые отвечают разрядами на каждое изменение ситуации, указывает на то, что в его основе лежит действие определенных нервных приборов.
С другой стороны, ориентировочный рефлекс обнаруживает ряд признаков, существенно отличающих его от обычных безусловных рефлексов: при неоднократном повторении одного и того же раздражителя явления ориентировочного рефлекса скоро угасают, организм привыкает к этому раздражителю, и его предъявления перестают вызывать описанные реакции. Это исчезновение ориентировочных реакций на повторяющиеся раздражители называется привыканием (habituation).[3]
Следует отметить, что это исчезновение ориентировочного рефлекса по мере привыкания может быть временным явлением, и достаточно небольшого изменения в раздражителе, чтобы ориентировочная реакция снова возникла. Это явление возникновения ориентировочного рефлекса при малейшем изменении раздражения иногда называется реакцией «пробуждения» (или arousal). Характерно, что такое появление ориентировочного рефлекса, как уже отмечалось выше, может иметь место не только при усилении, но и при ослаблении привычного раздражителя и даже при его полном исчезновении. Так, достаточно сначала «угасить» ориентировочные рефлексы на ритмически предъявляемые раздражители, а затем, после того как ориентировочные реакции на каждое раздражение угасли в результате привыкания, пропустить один из ритмически предъявляемых раздражителей. В этом случае отсутствие ожидаемого раздражителя вызовет появление ориентировочного рефлекса.
Всеми этими признаками своей динамики ориентировочный рефлекс существенно отличается от безусловного рефлекса. Следует отметить и тот факт, что ориентировочный рефлекс может быть вызван и условным раздражителем: его можно получить, если предъявлять животному условный сигнал, который будет говорить о появлении какого-либо изменения в окружающей обстановке. У человека таким сигналом может быть слово, которое легко вызывает у него явления подготовки, настораживания, ожидания появления сигнала и т. п.
Было бы неверным думать, что ориентировочный рефлекс носит характер общей, генерализированной активации организма. На самом деле он может иметь дифференцированный, избирательный характер, причем эта избирательность может проявляться как по отношению к возникающим сигналам, так и по характеру той готовности эффекторных двигательных аппаратов, которые вызываются «настороженностью».
Это легко видеть, если длительно предъявлять испытуемому какой-нибудь один сигнал, например звук определенной высоты, тогда в силу привыкания все реакции на этот звук будут угашены, однако это «привыкание» будет носить избирательный характер, и стоит только минимально изменить высоту звука, чтобы весь комплекс ориентировочных реакций снова появился. Такой прием позволил советскому исследователю Е. II. Соколову объективно оценить ту избирательность, которой характеризуются ориентировочные реакции («реакции пробуждения») в отношении дифференцированных сигналов, и говорить о «нервной модели стимула», которая обнаруживается с помощью этого приема.
Виды внимания
В психологии различают два основных вида внимания — непроизвольное и произвольное.
О непроизвольном внимании говорят в тех случаях, когда внимание человека непосредственно привлекается либо сильным, либо новым, либо интересным (соответствующим потребности) раздражителем. Именно с таким видом внимания мы имеем дело, когда помимо желания поворачиваем голову, если в комнате внезапно раздается стук, настораживаемся, когда слышим непонятные шумы или когда наше внимание привлекается каким-либо новым, неожиданным изменением обстановки.
Механизмы непроизвольного внимания у нас с животными общие. Мы уже говорили во вводной части главы о факторах этого вида внимания и о его нейрофизиологических основах, когда разбирали механизмы ориентировочного рефлекса. Легко видеть, что этот вид внимания имеет место уже у маленького ребенка, и следует лишь отметить, что на первых этапах оно носит неустойчивый и относительно узкий по объему характер (ребенок раннего и дошкольного возраста очень быстро теряет внимание к возникшему новому раздражителю, ориентировочный рефлекс у него быстро угасает или тормозится появлением любого другого раздражителя), объем его внимания относительно узок, и он не может распределить свое внимание между несколькими раздражителями, возвращаясь к предшествующему и не упуская из поля своего внимания предыдущий.[3]
Произвольное внимание свойственно лишь человеку. Оно долгое время оставалось загадкой для психологии, и на нем следует остановиться особо.
Основной факт, указывающий на наличие у человека особого типа внимания не свойственного животным, состоит в том, что человек произвольно может сосредоточивать свое внимание то на одном, то на другом объекте, даже в тех случаях когда в окружающей его обстановке ничего не меняется.
Наиболее известный пример произвольного внимания был дан французским психологом Рево д’Аллонном, он стал основой для его идеалистической философии. Если мы предложим человеку внимательно разглядывать шахматную доску, клетки которой сохраняют неизменный характер, то он соответственно нашей или собственной инструкции легко сможет выделить в этом однородном фоне самые разнообразные фигуры. В однородном и неменяющемся поле оказывается скрыто много разно образных структур, и человек по своему желанию может выделить любые новые структуры из этого неизменного поля. Иногда эта возможность произвольно выделять нужную структуру из поля появляется еще отчетливее, и соответственно своему желанию человек может выделять менее четкую структуру из более четких, преодолевая те законы структурного восприятия, которые описаны выше.
Таким образом, становится ясно, что человек может выходить за пределы естественных законов восприятия, не подчиняясь действию однородного фона или сильных перцепторных структур, а выделяя нужные ему структуры и меняя их по своему желанию.
Все эти факты дали Рево д’Аллонну основание для обоснования идеалистического представления о психических процессах человека, указывая, что если поведение животного подчиняется прямому воздействию среды, то поведение человека располагает возможностью произвольно распознавать любые схемы и подчинять свое поведение этой свободной «схематизации», которую он считал основным свойством человеческого духа.
Аналогичные явления можно было наблюдать и в организации движений человека; достаточно человеку решить, что он поднимет руку, чтобы его рука как 6ы автоматически поднялась; это явление известный психолог У. Джемс обозначил латинским термином «fiat!» (Да будет!), видя в нем самое простое доказательств существования свободной воли, которая не подчинялась законам природы, но сам определяла поведение человека.
Дальнейшие наблюдения показали, что даже простая мысль о предстоящем движении руки вызывает в ней отчетливое напряжение, которое можно зарегистрировать в изменении электромиограммы руки. Эти явления получили в психологии название «идеомоторных актов» и нередко приводились как иллюстрации или влияний представления на движение.
Наконец, такие же явления произвольного внимания можно наблюдать и в интеллектуальной деятельности, когда человек сам ставит перед собой соответствующую задачу и эта задача определяет дальнейшее избирательное течение ею ассоциаций.
Вот почему факты произвольного внимания относились в классических руководствах по психологии к разделу «Воля» и использовались для иллюстрации положения психики о том, что человек не подчиняется объективным законам природы, но зависит от влияний, исходящих из свободного духа.
Легко видеть, что все эти наблюдения описывали реально существующие факты, но объяснение этих фактов в рамках традиционной естественнонаучной психологии оказывалось невозможным, и именно это широко раскрываю двери для ненаучных идеалистических гипотез о влиянии «свободной воли — на протекание психических процессов человека.
Тупик, в который пришли попытки объяснить явления произвольного внимания в классической естественнонаучной психологии, можно преодолеть, если изменить традиционные представления о сознательных процессах, перестать рассматривать их как первичные, всегда существующие особенности духовной жизни и рассматривать их как продукт сложного общественно-исторического развития. Только сделав этот шаг и рассмотрев вопрос о генезе произвольного внимания, можно увидеть корни и подойти к его научному объяснению. Ребенок живет в окружении взрослых и развивается в процессе живого общения с ними.
Это общение, осуществляемое с помощью речи, действий и жестов взрослого, существенно влияет на организацию его психических процессов.
Ребенок раннего возраста рассматривает окружающую его привычную обстановку, и его взор блуждает по окружающим предметам, не останавливаясь ни на одном из них и не выделяя того или иного предмета из остальных. Мать говорит ребенку: «Это чашка!» и указывает на нее пальцем. Слово и указательный жест матери сразу же выделяют этот предмет из остальных, и ребенок фиксирует взглядом указанную чашку и тянется к ней рукой. В этом случае внимание ребенка продолжает носить непроизвольный внешне детерминированный характер, с той только разницей, что к естественным факторам внешней среды присоединяются факторы общественной организации его поведения, управление вниманием ребенка посредством указательного жеста и слова. В этом случае организация внимания разделена между двумя людьми: мать направляет внимание, ребенок подчиняется ее указательному жесту и слову.
Однако это является лишь первым этапом формирования произвольного внимания: внешним по источнику и социальным по природе. В процессе своего дальнейшего развития ребенок овладевает речью и оказывается в состоянии самостоятельно указывать на предметы и называть их. Развитие речи ребенка вносит коренную перестройку в управление его вниманием. Сейчас он уже оказывается в состоянии самостоятельно перемещать свое внимание, указывая на тот или иной предмет жестом или называя его соответствующим словом. Организация внимания, которая раньше была разделена между двумя людьми, матерью и ребенком, становится теперь новой формой внутренней организации внимания, социальной но своему происхождению, но внутренне опосредованной по своей структуре. Этот этап и следует считать этапом рождения новой формы произвольного внимания, которое оказывается не изначально присущей человеку формой проявления «свободного духа», а продуктом сложного социально-исторического развития.
На дальнейших этапах речь ребенка развивается: создаются все более сложные и подвижные внутренние речевые (интеллектуальные) структуры и внимание человека приобретает новые черты, становится управляемым внутренними интеллектуальными схемами, которые сами являются продуктом сложного социально го формирования психических процессов.
Все это показывает:
• что произвольное внимание человека с его подвижным и независимым oт непосредственных внешних воздействий характером действительно существует;
• имеет объяснимый детерминированный характер, является социальным по происхождению и опосредованным внутренними речевыми процессами по своему строению.
Мо мере дальнейшего развития внутренние речевые и интеллектуальные процессы ребенка становятся настолько сложными и автоматизированными, что перевод внимания с одного объекта на другой перестает требовать специальных усилий и приобретает характер той легкости и, казалось бы, «непроизвольности», которую ощущаем все мы, когда в нашем мышлении легко переходим с одного объекта на другой или когда оказываемся в состоянии длительно сохранять напряженное внимание по отношению к интересующей нас деятельности.
ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС — это… Что такое ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС?
- ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС
- (ориентировочная реакция) — сложная реакция организма на новый раздражитель, направленная на мобилизацию систем организма для выработки соответствующего действия в новой ситуации. О. р. лежит в основе ориентировочной деятельности человека, т. е. деятельности, направленной на обследование окружающих предметов в целях получения информации, необходимой для решения стоящих перед субъектом задач.
Поэтому О. р. называют также исследовательским рефлексом. В комплекс компонентов реакции О. р. входят: движение головы и глаз в направлении источника раздражителя, сосудистые изменения, изменение дыхания и электрического сопротивления кожи, возрастание тонуса мышц, а также повышение физиологической активности коры больших полушарий головного мозга, проявляющееся в форме уменьшения альфаритма ЭЭГ. Сенсорными компонентами реакции О. р. являются повышение чувствительности анализаторов и возрастание ритмической частоты слияния ощущений. Реакция О. р. возникает на новизну раздражителя. При предъявлении нового раздражителя вначале образуются все компоненты О. р. Через несколько его предъявлений часть компонентов О. р. угасает, а при последующих предъявлениях рефлекс угасает полностью. Раздражитель перестает быть новым для организма. Отличительная особенность угасания О. р.— его селективность по отношению к раздражителю- Изменяя разные параметры стимула, можно вновь вызвать реакцию О. р.
Энциклопедический словарь по психологии и педагогике.
2013.
- Ориентировочная реакция
- ОРОГЕНИТАЛЬНЫЙ СЕКС
Смотреть что такое «ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС» в других словарях:
ориентировочный рефлекс — сложная реакция животных и человека на новизну стимула, названная И. П. Павловым рефлексом «что такое?». Биологический смысл О. р. создание условий для лучшего восприятия раздражителя. Это достигается за счет появления комплекса соматических,… … Большая психологическая энциклопедия
Ориентировочный Рефлекс — комплекс реакций организма в ответ на новизну раздражителя, в который входят: движение головы и глаз в направлении источника раздражения, расширение сосудов мозга при одновременном сужении периферичес … Психологический словарь
Ориентировочный рефлекс — специфическая реакция животных, направленная на получение дополнительной информации путем создания условий для лучшего восприятия раздражителей. Выражается в общем прекращении текущей деятельности (стоп реакция), ориентации анализаторов на… … Словарь дрессировщика
ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС — См. ориентировочная реакция … Толковый словарь по психологии
рефлекс ориентировочный — комплекс реакций организма в ответ на новизну раздражителя сложная реакция животных и человека на новизну стимула, названная И. П. Павловым рефлексом «что такое?». Его биологический смысл создание условий для лучшего восприятия раздражителя. Это… … Большая психологическая энциклопедия
рефлекс безусловный — (рефлекс врожденный) наследственно закрепленная стереотипная форма реагирования на биологически значимые воздействия внешнего мира или на изменения внутренней среды организма. Рефлекс, всегда реализуемый при действии на организм определенных… … Большая психологическая энциклопедия
рефлекс врожденный — (рефлекс врожденный) наследственно закрепленная стереотипная форма реагирования на биологически значимые воздействия внешнего мира или на изменения внутренней среды организма. Рефлекс, всегда реализуемый при действии на организм определенных … Большая психологическая энциклопедия
РЕФЛЕКС ОРИЕНТИРОВОЧНЫЙ — сложная реакция животного или человека, вызываемая действием новых раздражителей или изменением старых, названная И.П. Павловым рефлексом «что та кое?». С Р. о. начинается любой сложнорефлекторный акт организма … Психомоторика: cловарь-справочник
Ориентировочная реакция — (рефлекс «Что такое?», по И. П. Павлову) комплекс сдвигов в разных системах организма животного или человека, вызываемый любым неожиданным изменением ситуации и обусловленный особой активностью центральной нервной системы. Изменения в… … Большая советская энциклопедия
ФОТОТАКСИС — Ориентировочный рефлекс на свет. Положительный фототаксис – движение по направлению к источнику света, отрицательный – от него. Этот термин предназначен для обозначения таких движений у животных; в отношении растений используется термин… … Толковый словарь по психологии
Ориентирующий рефлекс | Статья о Ориентирующем рефлексе по The Free Dictionary
(также фокусирующий рефлекс, исследовательский рефлекс, рефлекс Павлова «Что это такое?»), У животных и человека, комплекс реакций в различных системах организма, вызванный любым неожиданным изменением ситуации и обусловлено особой деятельностью центральной нервной системы.
Изменения в функционировании центральной и вегетативной нервных систем во время ориентировочного рефлекса направлены на мобилизацию анализирующей и двигательной систем организма, что способствует быстрой и точной оценке новой ситуации и выработке нового, неавтоматического действия.Подавление предшествующей активности и поворот головы (ушей, глаз) на раздражитель происходят одновременно. Ориентирующий рефлекс сопровождается повышением уровня адреналина в крови, изменением электрического потенциала кожи (кожный гальванический рефлекс), реакцией активации (в виде десинхронизации медленной электрической активности коры головного мозга) и ряд других явлений, характеризующих подготовку организма к действиям в новой ситуации. Функции, не участвующие в таких действиях, например пищеварение, подавляются.Когда изменение обстановки сопровождается безусловным раздражителем, т. Е. Когда оно усилено раздражителем, на основе ориентировочного рефлекса может развиться условный рефлекс; индифферентный раздражитель становится существенным и значимым для организма. Если нейтральный стимул повторяется, ориентировочный рефлекс больше не возникает.
Ориентирующий рефлекс играет важную роль в организации высшей нервной деятельности у животных и человека. Согласно современным представлениям, в основе ориентировочной реакции лежат активирующие воздействия ретикулярной формации на высшие отделы центральной нервной системы.Вместе с тем существенно повышается уровень возбудимости соответствующих зон коры больших полушарий, что создает благоприятные условия для формирования в коре условно-рефлекторной цепи.
У людей ориентировочный рефлекс участвует в актах различной степени сложности, от реакции на любой новый агент до самой сложной умственной работы, например, когда человек, столкнувшись с неожиданным фактом или мыслью, концентрируется и мобилизует себя на понять это.В основе возникающего в данной ситуации внимания лежит ориентировочная реакция, которая, по мнению В. М. Бехтерева, проявляется в виде «рефлекса концентрации».
Роль ориентировочной реакции в психической деятельности человека более полно раскрывается при нарушении этой активности, например, при шизофрении. Утрата ориентировочного рефлекса или его угасание при повторных раздражениях значительно снижает возможность адаптации к новым условиям. В других случаях наличие тормозящей составляющей ориентировочного рефлекса и отсутствие следственной составляющей не позволяет проанализировать новую ситуацию и адекватно на нее отреагировать.
СПИСОК ЛИТЕРАТУРЫ
Павлов И.П. Полное собрание сочинений , 2-е изд., Т. 3, книги 1–2. Москва-Ленинград, 1951.
Ориентировочный рефлекс и ориентировочно-исследовательская деятельность . Москва, 1958.
Магун, Х. Бодрствующий мозг . М., 1961. (Пер. С англ.)
Chauvin, R. Povedenie zivotnykh , Chapter 6. Москва, 1972. (Пер. С французского)
.
Attention — Scholarpedia
Рис. 1: Динамика экзогенной (красный) и эндогенной (зеленый) ориентации внимания. Пунктирная красная линия указывает на постоянное внимание к прогнозирующим или интересным прямым подсказкам.
Внимание относится к процессу, с помощью которого организмы выбирают подмножество доступной информации, на которой следует сосредоточиться для улучшенной обработки (часто в смысле отношения сигнал / шум) и интеграции. Обычно считается, что внимание имеет как минимум три аспекта: ориентацию, фильтрацию и поиск, и может быть сосредоточено на одном источнике информации или разделено между несколькими.Каждый из этих аспектов имеет определенные свойства, которые кратко обсуждаются ниже. Внимание и сознание тесно связаны, хотя эти два понятия можно различить как концептуально, так и эмпирически.
Ориентирование
Самый простой способ выбрать один из нескольких входных стимулов — это ориентировать наши сенсорные рецепторы на один набор стимулов и подальше от другого. Обычно мы не видим и слышим пассивно, мы активно смотрим или слушаем, чтобы видеть и слышать.
Ориентирующий рефлекс
Прототипом ориентирования является реакция собаки или кошки на внезапный звук.Животное быстро настраивает свои органы чувств, настораживая уши и поворачивая глаза, голову и / или тело, чтобы оно могло оптимально воспринимать информацию о событии. Такие реакции, как движение глаза в направлении звука или периферического движения, а также сопутствующие корректировки позы, изменения проводимости кожи, расширение зрачка, снижение частоты сердечных сокращений, пауза в дыхании и сужение периферических кровеносных сосудов происходят автоматически. и в совокупности именуются ориентирующим рефлексом .Наиболее эффективными ориентирующими стимулами являются громкие звуки, внезапно появляющийся яркий свет, изменения контуров или движения в периферическом поле зрения, которые не являются регулярными и предсказуемыми. Это как если бы у нас была внутренняя «модель» непосредственного мира стимулов вокруг нас. Когда мы замечаем отклонение входящего стимула от этой модели, мы рефлекторно ориентируемся на этот стимул, чтобы обновить эту модель как можно быстрее (Соколов, 1975). Если один и тот же стимул повторяется неоднократно, он становится ожидаемой частью нашей модели мира, и наш ориентировочный рефлекс на него становится слабее, даже если стимул достаточно сильный.Однако при изменении характера раздражителя рефлекс восстанавливается до полной силы.
Скрытое ориентирование
Открытая ориентировочная реакция на внезапные изменения в окружающей среде обычно сопровождается другой, невидимой ориентировочной реакцией, фиксацией внимания на событии или объекте, вызвавшем рефлекс. Это незримое ориентирование внимания называется скрытым ориентированием . Комбинация явной и скрытой ориентации на событие обычно приводит к усиленному восприятию этого события, в том числе к более быстрой идентификации и осознанию его значимости: например, мы можем переместить взгляд на объект ( явное внимание ) или переключить внимание на него. не двигая глазами ( скрытое внимание, ).Хотя эта скрытая ориентация внимания обычно происходит в связи с открытой ориентацией, рефлексивной или произвольной, можно скрыто присутствовать на событии или стимуле, не делая никаких явных признаков того, что мы это делаем (например, Helmholtz, 1867/1925; Posner , 1980; Wright & Ward, 2008). Таким образом, скрытая ориентация внимания обычно изучается отдельно от явного ориентировочного поведения, хотя они, несомненно, тесно связаны. Премоторная теория внимания (например, Rizzolatti et al, 1994) предполагает, что эти системы внимания поддерживаются одними и теми же нейронными механизмами, хотя подробное изучение поведенческих и физиологических данных показывает, что, хотя эти два, вероятно, имеют общие нейронные механизмы, они не являются идентичны (см. обсуждения Corbetta & Shulman, 2002; Wright & Ward, 2008).
Управляемый стимулами, экзогенный, ориентирующий
Как и в случае с ориентировочным рефлексом, внезапные или интенсивные стимулы могут вызывать скрытую ориентацию, то есть привлекать внимание. Например, внезапно появляющиеся буквы на мониторе компьютера привлекают внимание и реагируют на них быстрее, чем на постепенно появляющиеся буквы (Jonides & Yantis, 1988; Yantis & Jonides, 1984). Если такой внезапный стимул (прямой сигнал ) появляется примерно за 100 мс до другого стимула (цель ) в том же пространственном местоположении, последний обрабатывается быстрее и точнее, чем если бы он появился в другом месте ( е.g., Müller & Humphreys, 1991), предположительно потому, что внимание было рефлексивно привлечено к пространственному расположению прямого сигнала. Внимание, которое рефлекторно захватывается таким образом, называется ориентированным экзогенно, стимулом. Внимание, ориентированное таким образом, управляемым стимулом, не остается ориентированным на привлекающее место надолго, однако, перемещаясь в другое место через 100-200 мс или около того, если только привлекающий стимул не требует усиленной обработки или не сигнализирует о высокой вероятности появления цели в это место.На рис. 1 (красным) показан типичный временной ход экзогенно ориентированного внимания в месте непрогнозирующего и неинтересного прямого сигнала. Внимание также обращается на резкое появление нового объекта восприятия, даже если его появление не сопровождается изменением яркости, но не изменением яркости, которое не связано с появлением нового объекта (Yantis & Hillstrom, 1994). Таким образом, внимание привлекается как к новому объекту восприятия, так и к его местоположению (Egeth & Yantis, 1997).Экзогенный захват внимания также происходит на слух и в контакте. Более того, прямая подсказка в одной из этих модальностей, например, звук, может ориентировать внимание на место, так что, когда цель в другой модальности, скажем, визуальный паттерн, встречается там, она также обрабатывается более быстро и точно (Wright & Ward, 2008).
Часть эффекта реплики при ориентировании, управляемой стимулами, по-видимому, вызвана остаточной сенсорной активностью самого прямого сигнала, которая также исчезает в течение 100-200 мс (Wright & Ward, 2008).Действительно, одновременное представление до 4 прямых сигналов на одном дисплее может вызывать эффекты сигналов во всех местах их расположения (Wright & Richard, 2003). Тем не менее, эффект реплики от одного отображения прямой реплики больше, чем эффект от нескольких отображений реплики, что указывает на то, что эффекты реплики от единственной прямой реплики возникают как из-за ориентации внимания, так и из остаточной сенсорной активации, тогда как эффекты от множественных отображений реплик возникают исключительно из остаточной сенсорная активация (Wright & Richard, 2003).
Нейрофизиология ориентированного на стимулы
Рисунок 2: Подкорковые зоны внимания мозга.
Рефлексивный захват внимания резкими или интенсивными раздражителями реализуется сетью областей мозга, которая включает в себя верхний бугорок, пульвинарное ядро таламуса (оба подкорковые, рис. 2) и заднюю теменную кору, а также области лобной части. кора головного мозга и, конечно же, различные сенсорные коры. Многие исследования показывают, что ранняя модель, в которой задняя теменная кора головного мозга отвлекает внимание от текущей цели / местоположения, верхний бугорок смещает его к новой цели / местоположению, а ядро пульвинара привлекает внимание к этому новому локусу, примерно верна ( Райт и Уорд, 2008 г.).Недавние визуальные исследования выявили более конкретную картину корковых частей этой сети, которые включают височно-теменное соединение и вентрально-лобную кору, в частности нижнюю лобную извилину и среднюю лобную извилину, в основном на правой стороне мозга. (Корбетта и Шульман, 2002). На рис. 3 красным цветом показана правосторонняя вентропариетальная сеть , участвующая в ориентировании на внезапно возникающий зрительный стимул, такой как прямая реплика или неверно указанная цель.Подкорковые области сети скрыты на этом виде.
Рисунок 3: Области и сети коркового внимания (стрелочные соединения). Вентропариетальная сеть (экзогенная ориентация) — красным, дорсопариетальная сеть (эндогенная ориентация) — зеленым, сенсорная кора — оранжевым, взаимодействие между локусами теменной коры — синим.
Целеустремленная или внутренняя ориентация
В дополнение к рефлексивному ориентированию, основанному на стимулах, мы также можем направить внимание на место в пространстве или на объект добровольно ( эндогенно ) целенаправленно, часто на основе сигнала, который говорит нам, куда смотреть или для прослушивания, например, объявления по громкоговорителю о том, что пассажиры определенного рейса выйдут на посадку у определенных выходов в аэропорту.Информация о том, где или что смотреть или слушать, ожидая, когда произойдет экологическое событие, и мы часто готовимся к этому событию, ориентируя внимание на место и время ожидаемого события (LaBerge, 1995). Это продвинутое, целеустремленное выравнивание внимания улучшает обработку события, когда оно действительно происходит (например, Posner, 1980).
Стоимость и преимущества символических реплик
Ориентирование на цель обычно изучается в экспериментах путем предъявления прогнозирующей символической подсказки (например, стрелки, указывающей на возможное целевое местоположение) относительно того, где в пространстве будет возникать целевой стимул, так называемая парадигма Познера (e.грамм. Познер 1980). Обычно (скажем, в 80% испытаний с указанием стрелки, что может составлять 80% или более от общего числа испытаний) цель действительно встречается там (так называемые испытания с действительной подсказкой), что дает субъекту стимул сосредоточить внимание на ней до того, как цель внешность. Иногда (в 20% испытаний с указанием стрелки, когда 80% действительны) цель встречается в месте, отличном от указанного (это называется испытаниями с недопустимой меткой). Наконец, в некоторых реализациях представлена третья группа испытаний (испытания с нейтральной репликой; часто 20% от общего числа испытаний или меньше) представлена нейтральная (в отношении местоположения цели) реплика.На этих испытаниях мишени случайным образом встречаются в возможных местах. В этих условиях цели в испытаниях с действительными сигналами реагируют быстрее, чем цели в нейтральных испытаниях (преимущество ориентации на целевое местоположение), тогда как цели в испытаниях с неверными сигналами реагируют медленнее, чем цели в нейтральных испытаниях ( стоимость ориентации не в том месте). Такая целенаправленная ориентация внимания медленнее, чем ориентированная на стимулы, обычно требуется около 300 мсек для достижения полной эффективности (Shepard & Müller, 1989; рис. 1, зеленая линия).Более того, целенаправленное внимание может поддерживаться в определенном месте в течение довольно длительных периодов времени, даже нескольких минут, тогда как внимание, ориентированное на стимулы, обычно преходяще, если целенаправленное внимание не вызывается прямым прогнозирующим сигналом или интересным целевым стимулом. Ориентирование на цель не является автоматическим и может быть прервано стимулом, привлекающим внимание (Müller & Rabbitt, 1989), хотя оно также может поддерживаться даже в присутствии такого стимула, если прогностическая ценность символического сигнала высока. достаточно (Янтис и Джонидес, 1990).
Нейрофизиология целевого ориентирования
Определенная сеть областей мозга поддерживает целенаправленную ориентацию внимания. Корковые отделы называются дорсопариетальной сетью , (Corbetta & Shulman, 2002). На рисунке 3 зеленым цветом показано правое полушарие этой двусторонней сети. Он включает в себя лобные области, особенно лобные поля глаза (FEF), которые, по-видимому, также участвуют в произвольной ориентации на слуховые и тактильные события (например, Shomstein & Yantis, 2004), теменные области, в частности, интрапариетальную борозду (IPS), так как а также соответствующие сенсорные области коры и подкорковые области (таламус и верхний холмик, рис. 2).Более того, эта сеть взаимодействует с вентропариетальной сетью (показана красным), вероятно, через соединения между TPJ и IPS (показаны синим). Возможно, что взаимодействия между областями головного мозга дорсопариетальной и вентропариетальной сетей опосредуются синхронизацией их активности на различных частотах, в том числе особенно в гамма (30-70 Гц) и альфа (8-14 Гц) диапазонах. (например, Doesburg и др., 2008; см. также Варела и др. 2001; Ward, 2003).
Запрет возврата
Когда внимание было привлечено к определенному месту или объекту восприятия, а затем оно уходит в другое место или объект, кажется, что ему не удается вернуться в исходное местоположение или объект в течение периода до 2 секунд (например.г., Познер и Коэн, 1984; Самосвал, Драйвер и Уивер, 1991). Это называется запретом возврата (IOR). Предполагается, что IOR способствует поиску информативных объектов или мест, и это происходит внутри и между визуальными, слуховыми и тактильными модальностями, как и внимание (например, Klein, 2000; Spence & Driver, 1998; Ward, 1994), и в младенцы в возрасте 6 месяцев (Rothbart, Posner, & Boylan, 1990). Хотя возможно, что IOR возникает из-за торможения, связанного с некоторыми двигательными процессами, такими как движения глаз или мануальные реакции (обзор премоторной теории в этом отношении см. В Wright & Ward, 2008), недавние данные показывают, что также возможно, что его нервная выражение возникает во время перцептивной или когнитивной обработки стимулов в ранее посещенных местах (например,г., Prime & Ward, 2006).
Фокус внимания
Когда внимание направлено на определенный локус в пространстве, оно называется фокусным или пространственным вниманием. Если он ориентирован на определенный объект, будь то зрительный, слуховой, соматосенсорный, запах или вкус, он называется объектно-ориентированным вниманием . Но независимо от того, идет ли речь о месте или объекте, степень сосредоточения внимания может контролироваться целенаправленно или стимулированным образом (см. Yantis & Serences, 2003, для обзора пространственного и объектного внимания. и аналогичные им корковые механизмы).Чем больше степень распространения внимания, тем менее эффективна обработка информации в этой области (например, Laberge & Brown, 1989). Более того, чем дальше стимул от центра обслуживаемой области, тем менее эффективна его обработка (например, Eriksen & St. James, 1986). Этот последний эффект иногда называют градиентом внимания .
Фильтрация
Внимание действует как фильтр, извлекая больше информации из обслуживаемых стимулов и подавляя извлечение информации из оставленных без внимания стимулов.Подавление может быть настолько сильным, что может вызвать то, что было названо слепотой по невнимательности , которая происходит при всех модальностях.
Феномен коктейльной вечеринки
На шумной вечеринке с коктейлями (или любой другой) фильтрация внимания неумолима. Люди слушают один разговор, мелодию и т. Д., А остальное отфильтровывают как «шум». Черри (1953) использовал затенение для изучения этого явления в слуховом внимании. В этой технике наблюдатель должен повторить вслух (затенять) один из двух непрерывных речевых потоков.Близкое затенение приводит к потере большей части информации в незатененном потоке, за исключением иногда сильных прямых сигналов, таких как собственное имя или громкий шум, который может вызвать ориентацию на этот поток. Однако незатененная информация попадает в кратковременную память и может быть вызвана, если затенение было прервано.
Невнимательность и слепота к изменениям
Аналог слухового затенения использовался для изучения визуальной фильтрации (Neisser & Becklin, 1975). Субъекты затеняли одну из двух перекрывающихся видеопрограмм с аналогичными результатами: большая часть информации в потоке без затенения не была замечена или запомнена.Как и слуховая фильтрация, визуальная фильтрация позволяет мало отфильтрованной информации произвести неизгладимое впечатление. Современное воспроизведение этого результата привело к еще более драматичным и противоречащим интуиции результатам, например, к невозможности заметить, как человек в костюме гориллы бьет себя в грудь и скачет во время теневого баскетбольного матча (Simons & Chabris, 1999). Даже весьма драматические стимулы в простых проявлениях могут пройти незамеченными, если внимание будет направлено на что-то другое. Это было названо слепотой по невнимательности (Mack & Rock, 1998).Связанное с этим явление слепота к изменениям (например, Rensink, 2002), в котором сцена и та же сцена с ее изменением представлены в течение коротких периодов, разделенных пустым интервалом. Пока внимание не обращается на измененный элемент, о нем не сообщается.
Нейрофизиология фильтрации
Фильтрация внимания, по-видимому, осуществляется за счет активности пульвинарного ядра таламуса (рис. 2A) по инструкциям других, вероятно, лобных, кортикальных областей, таких как лобные поля глаза (FEF; рис. 2B; LaBerge, 1995).Предполагается, что ядро пульвинара является подкорковой областью, отвечающей за привлечение внимания, и оно проявляет повышенную активность, когда внимание необходимо использовать для фильтрации отвлекающих стимулов (например, Corbetta и др., 1991; Laberge & Buchsbaum, 1990). Влияние внимания на конкретный стимул на нервную активность проявляется довольно рано в сенсорной обработке и включает как усиленную реакцию на наблюдаемый стимул, так и подавленную реакцию на другие, оставленные без внимания стимулы. Например, применительно к зрению Моран и Дезимон (1985) показали, что реакция в зрительной области V4 обезьян на один и тот же раздражитель в значительной степени зависит от того, принимали его во внимание или игнорировали.Соответствующие обслуживаемые стимулы в рецептивном поле нейрона V4 получали энергичный ответ, который резко снижался, когда тот же самый стимул приходилось игнорировать. Точно так же внимание к определенной особенности объектов в поле зрения генерирует более энергичные ответы нейронов обезьяны V4, настроенных на эти особенности, чем когда внимание направлено на другие особенности тех же объектов (см. Обзор Maunsell and Treue, 2006 ). Более того, синхронизация в гамма-диапазоне нейронных ответов от разных нейронов в V4 на один и тот же стимул увеличивается за счет избирательного внимания, тогда как синхронизация в альфа-диапазоне уменьшается, что, возможно, служит для усиления сигнала, генерируемого присутствующими стимулами, по сравнению с сигналом, генерируемым дистракторы (Fries et al, 2001).Хороший обзор этих механизмов, который все еще актуален, содержится в Kastener and Ungerleider (2000).
Разделенное внимание
Очень сложно разделить внимание между двумя (или более) источниками. Например, люди не могут легко слушать два одновременных аудиопотока или просматривать два перекрывающихся видео при обнаружении целевых событий в каждом, особенно когда два источника пространственно разделены. Иногда два аспекта одного объекта могут быть успешно рассмотрены, но если эти два аспекта характеризуют два пространственно разделенных объекта, производительность хуже в условиях разделенного внимания (Bonnel & Prinzmetal, 1998).Также легче разделить внимание между информационными потоками в двух различных сенсорных модальностях, таких как зрение и слух, но если задача сложнее, чем простое обнаружение случайных стимулов в этих каналах, производительность все равно хуже, чем при обращении только к одному каналу (например, , Bonnel & Hafter, 1998). Когда задача более сложная, только если задача в одной модальности, например набор текста опытным машинистом, может быть выполнена автоматически может быть разделено внимание без снижения производительности, и только тогда, если способы ответа одинаково различны (например.g., опытный машинист, набирающий текст — визуальный / ручной, давая словесный ответ всякий раз, когда они слышат свое имя в слуховом канале — слуховом / вербальном).
В поисках
Когда мы знаем, что ищем, но не знаем, где это найти, мы должны искать это. В этом поиске очень важно внимание, и поисковые эксперименты дали много информации о его механизмах.
Простой поиск против сложного
Когда наблюдателям предоставляется поле с элементами для поиска определенной цели, они могут выполнять поиск очень быстро и примерно за одно и то же время независимо от количества нецелевых элементов, если цель отличается от нецелевых. в единственной функции; это называется простой / всплывающий / параллельный поиск (рис. 4, зеленая линия).Однако, если поисковик должен обнаружить совокупность признаков, поиск будет медленным, и время нахождения цели линейно увеличивается с количеством нецелевых объектов; это называется сложным поиском / последовательным поиском (рис. 4, красная линия). Легкий поиск может быть выполнен путем простого обнаружения наличия активации в конкретной одной из карт признаков, созданных сенсорной обработкой (карты признаков представляют собой листы зрительной коры, в которых активность определенных нейронов сигнализирует о наличии определенной особенности, такой как наклонная линия, на сетчатке).Это можно сделать, не обращая внимания на какой-либо конкретный элемент в данной области. С другой стороны, сложный поиск, по-видимому, требует сосредоточения внимания на каждом элементе по очереди, что замедляет поиск и делает общее время поиска зависимым от количества нецелевых элементов (например, Woodman & Luck, 1999). Теория интеграции признаков (например, Treisman & Gelade, 1980) объясняет эти данные, предполагая, что связывание признаков вместе в воспринимаемый объект требует фокуса внимания на конкретном локусе.Однако это не вся история, потому что соединения простых функций иногда также приводят к очень быстрому поиску (например, Wolfe et al, 1989), и внимание может быть последовательно направлено на элементы при простом поиске и на отдельные функции при поиске сочетаний. (Ким и Кейв, 1995).
Рисунок 4: Идеальные функции поиска для сложного и / или управляемого поиска (красная линия) и простого и / или автоматического поиска (зеленая линия).
Автоматический поиск в сравнении с контролируемым
Большая практика в сложной поисковой задаче, которая всегда требует одного и того же ответа на данный стимул, постепенно превращает процесс с управляемого поиска, , в котором время поиска является функцией количества элементов в поисковом наборе, на автоматический поиск, , в котором время поиска примерно не зависит от количества элементов в поисковом наборе (например,г., Schneider & Shiffrin, 1977; Шиффрин и Шнайдер, 1977). Функции поиска для управляемого и автоматического поиска напоминают функции для простого и сложного поиска (рис. 4). Распространенное объяснение процесса автоматизации состоит в том, что внимание постепенно отвлекается от управления задачами с увеличением практики, пока процесс не требует только минимального количества и считается автоматическим. В автоматическом поиске ответы баллистичны, поэтому их трудно запретить, а также они плохо запоминаются.В результате таких автоматических реакций в повседневной жизни могут возникать многие упущения внимания (например, Reason, 1984). С другой стороны, автоматическая обработка позволяет лучше разделить внимание.
Теории внимания
Каждый из основных аспектов внимания, ориентации, фильтрации и поиска породил множество теорий как на психологическом, так и на неврологическом уровне. Ранние теории, сфокусированные на фильтрации, утверждали, что обработка информации в мозге была структурно ограниченной, с ранним фильтром, основанным на физических характеристиках, таких как местоположение и спектральный состав, который пропускал только несколько выбранных стимулов (например,г., Бродбент, 1958). Демонстрация того, что по крайней мере некоторая обработка выполняется даже на отклоненных каналах, привела к отклонению этих теорий раннего отбора в пользу теорий позднего отбора , которые
предположил, что вся сенсорная информация проходит предварительный анализ. Вместо этого узкое место обработки возникает непосредственно перед входом в долговременную память (например, Deutsch & Deutsch, 1963). Существуют физиологические данные в пользу обоих подходов, и, вероятно, оба применяются в разных обстоятельствах (например,г., Пашлер, 1996).
Неудачи разделенного внимания привели к мысли, что внимание — это ограниченный ресурс, который, будучи востребован одной задачей, недоступен для другой (например, Kahneman, 1973). Демонстрация того, что разделение времени между перцептивными или когнитивными задачами возможно, особенно если одна из них слишком усвоена или автоматическая, с другой стороны, привела к идее, что существует множество ресурсов внимания, и что они могут быть разделены между задачами при условии, что нет конфликты (e.г., Виккенс, 1984). Согласно этому подходу, центральные ресурсы (такие как кодирование, сравнение, запоминание) взаимодействуют с пространственными и вербальными кодами, сенсорными модальностями и системами реакции, чтобы ограничить производительность. Этот подход подвергался критике как слишком гибкий (например, Navon, 1984), хотя идея ограниченных ресурсов внимания по-прежнему широко используется.
Другой класс теорий пытается описать механизмы ориентации внимания. Некоторые из них уже упоминались ранее.Подходы Корбетты и Шульман (например, 2002 г.), обсуждавшиеся выше (см. Рис. 3), Лаберже (например, 1995 г.) и Шиппа (например, 2004 г.), являются одними из наиболее общих и полезных. Многие из таких теорий подчеркивают концепцию значимости , , то есть ориентацию внимания на наиболее заметные из доступных мест или объектов, при этом заметность представляет собой сочетание восходящего и нисходящего вкладов. Шипп (2004) обсуждает несколько моделей значимости и объединяет их все в теории физиологии ориентирующей системы с картой заметности в пульвинарном ядре таламуса, объединяющей входные данные от других таких карт по всему мозгу.В этой теории IOR объясняется уменьшением заметности местоположения или объекта, которые недавно были проверены. См. Saliency_Map & Visual_Salience
Наконец, существуют другие модели, которые пытаются уловить определенные аспекты внимания в вычислительной или математической среде. Эпизодическая теория Сперлинга и Вайхзельгартнера (1995) носит особенно общий характер и учитывает множество возможных механизмов. В этом подходе предполагаются общие переходные функции времени для движения внимания, его появления и смещения в определенных местах.Эти временные функции относятся к прожектору внимания , который может иметь множество характеристик, таких как протяженность и интенсивность. Таким образом, теория может включать в себя множество результатов исследований всех аспектов, обсуждавшихся ранее. Совсем недавно Тейлор (например, Taylor & Rodgers, 2002, CODAM Model) предложил модель управления движением внимания, которая выражается в нейронной сети и является нейрофизиологически реалистичной. Существуют также теории колебаний внимания и увлечения фокуса внимания ритмическими событиями, такими как музыка (например,г., Large & Jones, 1999). Эти динамические теории также являются математическими, поскольку описание осцилляторов. Ни одна из этих теорий пока не получила всеобщего признания. Перейдите по ссылкам ниже, чтобы перейти к обсуждению нескольких конкретных теорий.
Внимание и сознание
Рисунок 5: Две модели отношений между вниманием и сознанием.
Внимание тесно связано с сознанием. Оба являются интегративными и в то же время избирательными. Невнимательная слепота, по-видимому, указывает на то, что предметы, не находящиеся во внимании, не воспринимаются сознательно.В самом деле, эта взаимосвязь настолько близка, что модель внимания Тейлора CODAM, как утверждается, также обеспечивает через последующий разряд сигналов контроля внимания два важных аспекта сознания, а именно. чувство владения сознательным опытом и его невосприимчивость к ошибкам, возникающим в результате неправильной идентификации. Тем не менее, эти две концепции можно различить. Внимание обычно концептуализируется как повышение отношения сигнал / шум как за счет подавления обработки необслуживаемых стимулов, так и за счет улучшенной обработки обслуживаемых стимулов.Сознание относится в первую очередь к самому феноменальному опыту и, во вторую очередь, к аспектам этого опыта, таким как его целостность, его чувство принадлежности к себе (онтогенез от первого лица), способность сообщать о своем содержании устно или другими способами, а также осознание бытия. сознание (метасознание). Существует несколько моделей взаимоотношений между вниманием и первичным, или феноменальным, сознанием, включая модель, предложенную Тейлором и большинством других, в которых когнитивный материал (ощущения, восприятия, познания, воспоминания и т.) либо обслуживаются, либо остаются без присмотра, при этом обслуживаемые пункты проверяются, а также подлежат отчетности и т. д. (рис. 5, вверху). В одной конкурирующей модели, среди прочих, весь когнитивный материал либо сознательный, либо бессознательный, при этом внимание отбирает часть сознательного материала для расширенной обработки, что делает его отчетным (рисунок 5, внизу; например, Lamme, 2003). Модель Ламме предполагает, что вся реентерабельная нейронная обработка порождает сознательный опыт, но только опыт, выбранный вниманием, может быть передан в отчет.Это означает, что сознание может существовать без внимания, но может быть обработан только сознательный материал. Напротив, Кох и Цучия (2007) приводят доказательства более полного разделения между ними, то есть внимания как с сознанием, так и без него, и сознания как с вниманием, так и без него. Согласно этой последней точке зрения, внимание и сознание — это отдельные процессы в мозге, но они тесно связаны. Пока нет однозначного выбора среди этих моделей.
Список литературы
- Боннель А.М. Хафтер Э. (1998) Разделил внимание между одновременными слуховыми и визуальными сигналами. Percep. Psychophys. 60: 179—190.
- Bonnel A-M Prinzmetal W. (1998) Разделение внимания между цветом и формой предметов. Percep. Psychophys. 60: 113-124.
- Broadbent DE. (1958) Восприятие и общение. Нью-Йорк: Pergamon Press.
- Черри К. (1953) Некоторые эксперименты по распознаванию речи одним и двумя ушами.J. Acoust. Soc. Amer. 25: 975-979.
- Corbetta M Miezin F Dobmeyer S Shulman G Petersen S. (1991) Выборочное и разделенное внимание во время визуального различения формы, цвета и скорости: Функциональная анатомия с помощью позитронно-эмиссионной томографии. J. Neurosci. 11: 2383-2402.
- Корбетта М Шульман GL. (2002) Контроль целенаправленного и стимулированного внимания в мозге. Nature Rev. Neurosci. 3: 201-215.
- Deutsch JA Deutsch D. (1963) Внимание: некоторые теоретические соображения.Psychol. Откр. 70: 80—90.
- Doesburg SM Roggeveen AB Kitajo K Ward LM (2008) Крупномасштабная фазовая синхронизация в гамма-диапазоне и избирательное внимание. Cereb. Cortex 18: 386-396.
- Egeth HE Yantis S (1997) Визуальное внимание: контроль, представление и ход времени. Энн. Rev. Psychol. 48: 269—297.
- Фрис П. Рейнольдс, Дж. Х. Рори, А. Е. Дезимон, Р. (2001) Модуляция колебательной нейронной синхронности посредством избирательного внимания. Наука 291: 1560-1563.
- Гельмгольц Х. фон (1867/1925) Трактат по физиологической оптике (Пер.Из 3-го немецкого издания). Нью-Йорк: Дувр.
- Йонидес Дж. Янтис С. (1988) Уникальность внезапного начала в привлечении внимания. Percep. Psychophys. 43: 346-354.
- Канеман Д. (1973) Внимание и усилия. Энглвуд Клиффс, Нью-Джерси: Прентис-Холл.
- Kastner S Ungerleider LG (2000) Механизмы зрительного внимания в коре головного мозга человека. Энн. Rev. Neurosci. 23: 315-341.
- Klein RE. (2000) Запрет на возвращение. Тенденции Cog. Sci. 4: 138-147.
- Koch C Tsuchiya N (2007) Внимание и сознание: два различных мозговых процесса.Тенденции Cog. Sci. 11: 16-22.
- ЛаБерж Д. (1995) Обработка внимания: искусство внимательности мозга. Кембридж, Массачусетс: Издательство Гарвардского университета.
- LaBerge D Buchsbaum MS (1990) Позитронно-эмиссионные томографические измерения активности пульвинара во время задания на концентрацию внимания. J. Neurosci. 10: 613-619.
- Ламме ВАФ. (2003). Почему различаются визуальное внимание и осведомленность. Тенденции Cog. Sci. 7: 12—18.
- Large EW Jones MR (1999) Динамика посещения: как люди отслеживают изменяющиеся во времени события.Psychol. Откр. 106: 119–159.
- Mack A Rock I. (1998) Невнимательная слепота. Кембридж, Массачусетс: MIT Press.
- Maunsell JHR Treue S (2006) Внимание, основанное на особенностях зрительной коры. Тенденции в Neurosci. 29: 317-322.
- Moran J Desimone R (1985) Селективное внимание контролирует визуальную обработку в экстрастриальной коре головного мозга. Наука 229: 782-784.
- Müller HJ Humphreys GW. (1991) Обнаружение приращения яркости: ограниченная мощность или нет? J. Exp. Психол .: Гум. Percep. Perfor.17: 107-124.
- Навон Д. (1984) Ресурсы: Теоретический суповой камень? Psychol. Откр.91: 216—234
- Neisser U Becklin R. (1975) Выборочный взгляд: внимание к визуально определенным событиям. Cog. Psychol. 7: 480-494.
- Пашлер Х. (1996) Психология внимания. Кембридж, Массачусетс: MIT Press.
- Posner MI. (1980) Ориентация внимания. Quart. J. Exp. Psychol. 32: 3-25.
- Познер М.И. Коэн Ю. (1984) Компоненты зрительного внимания. В H. Bouma DG Bouhuis (ред.), Внимание и производительность X (531–556). Хиллсдейл, Нью-Джерси: Эрлбаум.
- Prime DJ Ward LM. (2006) Корковые выражения торможения возврата. Brain Res. 1072: 161-174.
- Reason J. (1984) Отсутствие внимания в повседневной жизни. В R Parasuramen Д. Р. Дэвис (ред.) Разновидности внимания (515-549). Орландо: академический.
- Ренсинк РА. (2002) Обнаружение изменений. Энн. Rev. Psychol. 53: 245—277.
- Rizzolatti G Riggio L Sheliga BM (1994) Пространство и избирательное внимание.В C Umilta M Moscovitch (ред.) Внимание и исполнение XV (231-265). Кембридж, Массачусетс: MIT Press.
- Ротбарт М.К. Познер М.И., Бойлан А. (1990) Регулирующие механизмы в развитии младенца. В JT Enns (Ed) Развитие внимания: исследования и теория (47-66). Амстердам: Эльзевир.
- Шнайдер Шиффрин РМ. (1977) Управляемая и автоматическая обработка информации человеком I: Обнаружение, поиск и внимание. Psychol. Откр. 84: 1-66.
- Simons DJ Chabris CF. (1999) Гориллы среди нас: Устойчивая невнимательная слепота к динамическим событиям.Perception 28: 1059-1074.
- Шиффрин Р.М. Шнайдер В. (1977) Управляемая и автоматическая обработка информации человеком II: Восприятие обучения, автоматическое посещение и общая теория. Psychol. Откр. 84: 127-190.
- Шипп С. (2004) Схема внимания мозга. Тенденции Cog. Sci. 8: 223-230.
- Соколов Е.Н. (1975) Нейронные механизмы ориентировочного рефлекса. Соколов, О. С. Виноградова (ред.), Нейронные механизмы ориентирующего рефлекса (217–238). Нью-Йорк: Вили.
- Sperling G Weichselgartner E. (1995) Эпизодическая теория динамики пространственного внимания. Psychol. Ред. 102: 503-532.
- Спенс С. Ллойд Д. МакГлоун Ф. Николлс М.Э. Драйвер Дж. (2000) Запрещение возврата является супрамодальным: демонстрация всех возможных сочетаний зрения, осязания и прослушивания. Exp. Brain Res. 134: 42-48.
- Тейлор Дж. Г. и Роджерс М. (2002) Модель управления движением внимания. Нейронные сети 15: 309-326
- Самосвал SP Driver J Weaver B.(1991) Объектно-ориентированное торможение возвращения зрительного внимания. Quart. J. Exp. Psychol. А 43: 289–298.
- Treisman AM & Gelade G. (1980) Теория внимания с интеграцией признаков. Когнитивная психология, 12 (1), 97-136.
- Варела Ф. Лашо Дж. П. Родригес Е. Мартинери Дж. (2001) Сеть мозгов: фазовая синхронизация и крупномасштабная интеграция. Туземный Rev. Neurosci. 2: 229—239.
- Ward LM. (1994) Супрамодальные и специфические для модальности механизмы сдвигов слухового и зрительного внимания, вызванных стимулами.Жестяная банка. J. Exp. Psychol. 48: 242-259.
- Ward LM. (2003) Синхронные нейронные колебания и когнитивные процессы. Тенденции Cog. Sci. 7: 553—559.
- Виккенс CD. (1984) Обработка ресурсов во внимании. В R Parasuramen DR Дэвис (ред.) Разновидности внимания (63-101). Орландо: академический.
- Wolfe JM Cave KR Franzel SL. (1989) Управляемый поиск: альтернатива модели интеграции функций для визуального поиска. J. Exp. Психол .: Гум. Percep. Выполнить. 15: 419—433.
- Вудман GF Luck SJ.(1999) Электрофизиологическое измерение быстрых перемещений внимания во время визуального поиска. Природа 400: 867–869.
- Райт РД Ричард СМ. (2003) Сенсорное посредничество при захвате внимания на основе стимулов при отображении нескольких сигналов. Percep. Psychophys. 65: 925—938.
- Райт Р. Д. Уорд Л. М.. (2008) Ориентация внимания. Нью-Йорк: Издательство Оксфордского университета.
- Yantis S Hillstrom AP. (1994) Захват внимания, управляемый стимулами: свидетельство равносилюминантных визуальных объектов. J. Exp.Психол .: Гум. Percep. Выполнить. 20: 95-107.
- Янтис С. Йонидес Дж. (1984) Резкие визуальные проявления и избирательное внимание: данные визуального поиска. J. Exp. Психол .: Гум. Percep. Выполнить. 10: 601-621.
- Янтис С. Серенс JT (2003) Корковые механизмы космического и объектного контроля внимания. Тек. ОПИН. Biol. 13: 187-193.
Внутренние ссылки
- Валентино Брайтенберг (2007) Мозг. Scholarpedia, 2 (11): 2918.
- Джон Г.Тейлор (2007) Модель CODAM. Scholarpedia, 2 (11): 1598.
- Кейт Рейнер и Моника Кастельхано (2007) Движения глаз. Scholarpedia, 2 (10): 3649.
- Говард Эйхенбаум (2008) Память. Scholarpedia, 3 (3): 1747.
- Родольфо Ллинас (2008) Нейрон. Scholarpedia, 3 (8): 1490.
- Джефф Моелис, Кресимир Йосич, Эрик Т. Ши-Браун (2006) Периодическая орбита. Академия наук, 1 (7): 1358.
- Джон Доулинг (2007) Retina.Scholarpedia, 2 (12): 3487.
- Эрнест Баррето (2008) Shadowing. Scholarpedia, 3 (1): 2243.
- Аркадий Пиковский и Майкл Розенблюм (2007) Синхронизация. Академия наук, 2 (12): 1459.
- С. Мюррей Шерман (2006) Таламус. Академия наук, 1 (9): 1583.
Рекомендуемая литература
- Нятянен Р. (1992) Внимание и функции мозга. Махва, Нью-Джерси: Лоуренс Эрлбаум.
- Парасураман Р. (2000) Внимательный мозг.Кембридж, Массачусетс: MIT Press.
- Пашлер HE. (1996) Психология внимания. Кембридж, Массачусетс: MIT Press.
- Posner MI. (Эд) (2004) Когнитивная неврология внимания. Нью-Йорк: Guilford Press.
- Спенс С. Драйвер Дж. (Редакторы) (2004) Crossmodal Space and Crossmodal Attention. Нью-Йорк: Издательство Оксфордского университета.
- Райт Р. Д. Уорд Л. М.. (2008) Ориентация внимания. Нью-Йорк: Издательство Оксфордского университета.
Внешние ссылки
См. Также
МОДЕЛЬ КОДАМА, Сознание, Внимание и Сознание, Возбуждение, Настороженность, Визуальное внимание, Предвзятая модель конкуренции внимания, Моргание внимания, Невидимая слепота, Карта значимости, Модели внимания
,
ПАВЛОВСКИЙ ОРИЕНТИРУЮЩИЙ РЕФЛЕКС * — Скачать PDF бесплатно
Транскрипция
1 ACTA NEUROBIOL. IDSP. 1987, 47: ПАВЛОВСКИЙ ОРИЕНТИРУЮЩИЙ РЕФЛЕКС * Богуслав ЗЕРНИЦКИЙ Отделение нейрофизиологии, Институт экспериментальной биологии им. Ненцкого, Пастера 3, Варшава, Польша Ключевые слова: ориентировочный рефлекс, прицельный рефлекс, окулярный рефлекс фиксации Аннотация.Понятие ориентировочного рефлекса обсуждается с исторической точки зрения. Предлагаются следующие выводы: 1) в соответствии с духом концепции Павлова ориентировочный рефлекс — это реакция на неожиданный в данной ситуации раздражитель; (2) рефлекс состоит из компонентов прицеливания, возбуждения и восприятия; (3) рефлекс имеет три стадии, которые в целевом компоненте представлены движением к стимулу, поддержанием фиксации стимула и обратным движением; (4) когда рефлекс вызывается новым стимулом, обучение восприятию происходит на его второй стадии.Рассмотрены неориентирующие прицельные рефлексы. Предположительный котелок рассматривается как модель для исследования глазного ориентировочного рефлекса. Эта лекция дает мне возможность оглянуться на мои первые годы работы, когда под руководством Ежи Конорского я изучал слюнные условные рефлексы в павловской камере. Мой учитель, который ввел условнорефлекторный метод анализа двигательной активности собак, в свою очередь был учеником Павлова, в лаборатории которого он проработал два года в начале 1930-х годов.Идея ориентировочного рефлекса зародилась в лаборатории Павлова в начале нашего века. Павлов впервые упомянул об этом в лекции, прочитанной в Петербурге в 1910 году (13). Напомним * Лекцию, прочитанную на круглом столе «Анализ павловской обусловленности», посвященном 50-летию со дня смерти Павлова. Круглый стол проводился во время ежегодного собрания Европейского общества мозга и поведения в Марселе в сентябре 1986 года.
 in accordance with the spirit of Pavlov")
2 генезиса этой концепции.Установлено, что одним из факторов, нарушающих условные рефлексы слюны у собак, являются случайные раздражители (12, 13). Они были либо совершенно новыми стимулами для животного, либо новыми стимулами в данной ситуации. Таким образом, все они имели одну общую черту — быть неожиданными. Часто это были слуховые раздражители, в основном случайные звуки. Главным образом по этой причине Павлов построил свою знаменитую «тихую башню», где экспериментальные камеры звукоизолированы. Беспокоящий раздражитель этого типа вызывал характерное движение глаз, головы, а иногда и всего тела в направлении раздражителя, что явно облегчало восприятие раздражителя.Такой ответ Павлов назвал «рефлексом что это такое» и рефлексом ориентировочным. Каждый из этих терминов подчеркивал свою особенность, и ни один из них не был особенно подходящим, поскольку их буквальное значение было шире, чем описываемое явление (см. Ниже). Со временем получил признание второй срок. В лаборатории Павлова в основном интересовались ориентировочными рефлексами, вызываемыми указанными выше стимулами, лишенными определенного биологического значения, которые поэтому назывались «индифферентными». Однако ориентировочный рефлекс может быть вызван и неожиданным раздражителем, имеющим сильное биологическое действие.В таком случае могут возникнуть две ситуации. Во-первых, именно благодаря ориентировочной реакции раздражитель признается биологически значимым, и, таким образом, ориентировочный рефлекс составляет первую стадию цепного рефлекса. Например, благодаря ориентировочному рефлексу животное узнает хищника, и в результате возникает рефлекс бегства. Во второй ситуации стимул вызывает две реакции одновременно. Например, приложение болевого раздражителя к лапе животного вызывает одновременно с реакцией сгибания ориентировочную реакцию, заключающуюся в повороте головы в сторону раздражаемого места на коже.Примечательно, что в последнем случае ориентировочный рефлекс может нарушить течение своего неориентирующего аналога. Например, он может подавлять или растормаживать условный слюнный рефлекс. Это можно проиллюстрировать на примере лаборатории Павлова. После выработки слюнного условного рефлекса на определенный раздражитель применяется новый аналогичный раздражитель, который вызывает ориентировочную реакцию и не усиливается, чтобы вызвать условную дифференциацию. Этот раздражитель из-за явления генерализации также вызывает слюноотделение, но из-за тормозящего воздействия ориентировочного рефлекса он слабый на первом опыте.Однако, несмотря на отсутствие подкрепления, в следующих испытаниях слюноотделение постепенно увеличивается (Таблица I). Постепенное привыкание к ориентировочному рефлексу позволяет более полно проявить феномен генерализации. Этот процесс можно наблюдать, когда новый стимул применяется несколько раз за один экспериментальный сеанс

3 Освобождение генерализации условного слюнного рефлекса от внешнего торможения в процессе привыкания ориентировочного рефлекса.Реакция на новый, неусиленный раздражитель (квадрат) усиливается в течение трех дней подряд. Слюноотделение (в делениях шкалы) регистрировали в течение 30 с воздействия раздражителя. Выписка из эксперимента в лаборатории Павлова в (Из Павлова, 12). Временной стимул Подкрепление слюноотделения 28 ноябрь, круг 14 Да квадрат 3 нет ноя круг Да квадрат 7 нет ноя круг 15 Да квадрат 10 нет, и когда он применяется только один раз в каждой последовательной экспериментальной сессии (Таблица I). Только в ходе ряда испытаний постепенно угасает реакция слюны на неусиленный раздражитель.В лаборатории Павлова интерес был сосредоточен на двигательной, направленной составляющей ориентировочного рефлекса. В последующие годы, главным образом в первые десятилетия после Второй мировой войны, был описан ряд неспецифических элементов ориентировочного рефлекса: расширение зрачков, возбуждение ЭЭГ, психогальванический ответ, усиление мозгового кровотока и т. Д. Для этого комплекса реакций я предложите название «возбуждающий компонент» ориентировочного рефлекса. Следует подчеркнуть, что у него очень сильный ориентировочный рефлекс.Уровень возбуждения достаточен, чтобы подготовить животное к быстрой активности, если неожиданный стимул будет иметь биологическое значение. Интенсивность компонента возбуждения и легкость регистрации некоторых его элементов отвлекли внимание от направленного компонента ориентировочного рефлекса. Например, в некоторых исследованиях привыкания к ориентировочному рефлексу регистрировалось только возбуждение ЭЭГ. Более того, увлечение компонентом возбуждения, вероятно, является одной из причин, по которой многие считают ориентировочный рефлекс неспецифическим (см. 16, 17).Подчеркивается, что рефлекс вызывается стимулами любой модальности (зрительной, слуховой и т. Д.), А также появлением и исчезновением стимула. Однако направленная составляющая ориентировочного рефлекса определенно специфична. В то время как величина компонента возбуждения увеличивается с интенсивностью стимула, направление и величина компонента направления зависят от местоположения стимула. Например, сильный зрительный стимул появляется на маленьком
.")
На расстоянии
4 от точки фиксации глаза вызывает сильное возбуждение, но лишь небольшое движение глаз, которое точно переносит изображение стимула в центр сетчатки (26).Есть еще одна двусмысленность. Направленная реакция, способствующая восприятию стимула, очевидно, возникает не только на неожиданный стимул. По этой причине некоторые авторы (см. 2, 8) считают, что термин ориентировочный рефлекс должен иметь широкое значение. На мой взгляд, выход из этих концептуальных и терминологических трудностей был предложен Конорским в 1967 году, который использовал широкий термин «рефлекс прицеливания», определяемый как корректировка нижних уровней данной сенсорной системы для наилучшего восприятия стимула, главным образом благодаря действие подходящих эффекторов, связанных с данной рецептивной областью (6).Например, глазные прицельные рефлексы состоят в повороте глаз в направлении заданного объекта (рефлекс фиксации), их сближении и аккомодации. Используя этот термин и принимая во внимание первоначальное значение ориентировочного рефлекса, я предлагаю выделить в нем три составляющих: «прицельный», «возбуждающий» и «перцептивный». Рассмотрение последнего как рефлекторного компонента является произвольным, поскольку центральные процессы восприятия, по большей части, не являются прямым звеном открытой реакции.Предлагаю также различать в ориентировочном рефлексе три стадии. Поскольку они отчетливо видны в его целевом компоненте (рис. I), я назову их «этапом движения к стимулу», «этапом поддержания фиксации стимула» и «этапом обратного движения». Проанализируем некоторые свойства ориентировочного рефлекса в такой последовательности. Можно предположить, что раздражитель появляется на периферии рис. 1. Ориентирующий рефлекс фиксации у кошки. Стимул представлял собой колеблющуюся горизонтально горизонтальную световую щель (отмечена тонкой линией).Ордината: угол наклона глаза; нулевая точка относится к среднему положению глаза у кошки перед тройничком в состоянии покоя. Рефлекс регистрировался с правого глазного яблока, в то время как левое было закрыто. В положении покоя глаз был наклонен вверх примерно на 2 дюйма. При сохранении фиксации отклонение глаза вверх было примерно на 5 дюймов. Возвратное движение появлялось примерно через 1 с после исчезновения стимула. Рефлекс зарегистрирован у острой предтригеминальной кошки. (Из Зерницкого, 24).

5 сенсорное поле должно быть немедленно классифицировано как принадлежащее или нет, набор стимулов, ожидаемых в данной ситуации.Если раздражитель неожиданный, обязательно появится ориентировочный рефлекс, если ожидаемый, рефлекс не последует. Предположительно заторможены ориентировочные рефлексы на ожидаемые раздражители. В этом подавлении важную роль играет префронтальная кора. У префронтальных животных резко возрастает сопротивление привыканию ориентировочного рефлекса (23), и этот рефлекс может проявляться на условные раздражители (21). На втором этапе животное предположительно оценивает, известен ли уже неожиданный стимул, т.е.е. если он появился в его жизни. В соответствии с концепцией гностических единиц Конорского (6) и в некоторой степени с концепцией нейронных моделей Соколова (14, 17-20) можно предположить, что свойства стимула сравниваются с существующими единицами восприятия для знакомых стимулов. При отсутствии единицы восприятия может быть сформирована новая (см. 6). Прекращение этого перцептивного обучения может быть сигналом о том, что должно произойти обратное движение, даже если стимул все еще действует. Если тот же самый стимул появляется снова через короткое время, это в некоторой степени ожидаемо, а ориентировочный рефлекс слабее, часто даже наполовину.При повторении раздражителя усиливается процесс привыкания. Можно предположить, что перцептивная единица, хорошо сформировавшаяся благодаря повторению стимула, может быть активирована даже из периферической части сенсорного поля. Прицельные рефлексы, очевидно, выступают не только как составная часть ориентировочного рефлекса. Однако в условнорефлекторных опытах неориентирующие прицельные рефлексы наблюдаются нечасто. В павловских экспериментальных ситуациях некоторые собаки, прежде чем повернуть голову к миске с едой, ритуально смотрят на условный раздражитель, например, на движущийся метроном.Сильные целевые рефлексы типичны в экспериментах по инструментальному кондиционированию собак и кошек, если источник раздражителя находится далеко от места подачи пищи (4, 9). У птиц прицельный рефлекс возникает при классическом кондиционировании — голубь приближается и, наконец, клюет источник условного светового раздражителя, так называемый автоформный ответ (1, 2, 5). Наконец, инструментальная условная целенаправленная реакция может быть легко выработана пищевым подкреплением (15). Следует подчеркнуть, что неориентирующий целевой рефлекс обычно сопровождается лишь небольшим компонентом возбуждения, а не обучением восприятию.Кроме того, он контролируется определенным влечением (например, голодом), и поэтому он не приучается. Другими словами, его биологический смысл отличается от ориентировочного рефлекса. Неориентирующие рефлексы нацеливания могут проявляться длинными последовательностями. Примером может служить животное, наблюдающее за своим противником во время драки.
 and this reflex can appear to conditioned stimuli (21).")
6 Такие рефлексы часто представляют собой элемент сложного поведения, включающего также локомоторные и манипулятивные рефлексы.Они управляются определенной программой, управляемой, в свою очередь, определенным влечением (любопытство, голод, страх и т. Д.). Характерные длинные последовательности целевых рефлексов типичны для человеческого поведения. Как правило, они служат для заполнения хранилища памяти (например, при чтении книги) или имеют просто гедонистическую ценность (например, при взгляде на скульптуру, см. 22). Прицельные рефлексы регистрировать труднее, чем локомоторные и манипулятивные рефлексы. Особенно сложно изучить легко приучающийся целеуказательный компонент ориентировочных рефлексов.Однако в последнее время в этой области был достигнут методологический прогресс. Движение глаз, важный элемент многих рефлексов прицеливания, теперь можно точно регистрировать у свободно движущихся животных. В нашей лаборатории для изучения окулярно-фиксирующего компонента ориентировочного рефлекса мы использовали претригеминальную кошку, которую получают путем пересечения ствола мозга прямо перед корешками тройничных нервов (24). Хотя рефлекс фиксации менее точен, чем у интактного животного, его можно более удобно проанализировать в этом препарате (см. 24).Помимо прочего, его не беспокоят другие раздражители, причем отсутствие болевых раздражителей имеет особое значение. Предположительная подготовка — это, в некотором смысле, совершенная павловская «тихая башня», показанная на рис. 2. Ориентация глазных следящих рефлексов у острой предтригминальной кошки. Стимулом служила горизонтальная черная палка (1х4 ‘), движущаяся вверх по вертикальному меридиану или параллельно до 40 футов в носовом или височном полушарии. Усредненные четыре рефлекса для каждого локуса обозначены жирной линией, а представление палки — тонкой линией.Небольшие колебания в основном представляют собой шум записывающей системы. Маленькие числа обозначают порядок четырех серий рефлексов. Положение покоя глаза было постоянно низким, иногда даже ниже, чем положение начала стимула. Другие объяснения, как на рис. 1. (Из Michalski et al., 11).

7 Этот футляр адаптирован для изучения рефлекса фиксации глаза.Рефлекс, представленный на рис. 1, был зафиксирован в предтрубной подготовке. Обычно мы предъявляем стимул, который движется по полю зрения; он намного сильнее неподвижного. Если раздражитель движется медленно с постоянной скоростью, глазные яблоки в стадии фиксации следуют за ним аналогичным образом: появляется следующий рефлекс. На рис. 2 показано ориентирование по рефлексам, производимым маленьким предметом, движущимся вертикально через различные части поля зрения предтригминальной кошки. Из-за отсутствия горизонтальных движений глаз (их координационный центр находится кзади от перерезки ствола головного мозга) вертикальный рефлекс также вызывается стимулом, движущимся в боковом поле зрения.С помощью предтрубного препарата мы исследовали некоторые свойства фиксирующего компонента ориентировочного рефлекса у кошки (11, 25), а также влияние церебральных (см. 24) и «экологических» (7) поражений. На рисунке 3 показан пример наших результатов. Сдвиг отсутствия зрительного опыта Рис. 3. Сравнение свойств рефлекса ориентирования у нормально выращенных кошек (светлые кружки) и кошек, лишенных зрения в течение первых шести месяцев жизни (закрашенные кружки). Запись производилась на острых предтрубчатых препаратах.Стимулом служила горизонтальная черная палка (1×4 ‘) или световая щель того же размера, движущаяся вверх или вниз на разном расстоянии от вертикального меридиана. Средние данные для всех кошек и для четырех разновидностей стимулов (щель и палка, движение вверх и вниз). Для вертикального смещения следующих движений (последний график) отрицательные значения представляют отставание движения глаз относительно движения стимула. Другие объяснения, как на рис. 1. (Из Kossut et al., 7).

8 в молодом возрасте (кошки были лишены зрения в течение первых шести месяцев) нарушил следующий рефлекс.Его амплитуда и скорость уменьшаются, а задержка увеличивается. Кроме того, увеличивается смещение между положением изображения и вертикальным меридианом сетчатки. К этому следует добавить, что у кота до тройничного нерва удобно изучать аккомодационный компонент ориентировочного рефлекса (3) и некоторые элементы его возбуждающего компонента. СПИСОК ЛИТЕРАТУРЫ 1. БРАУН, П. Л. и ДЖЕНКИНС, Х. М. Автоматическое формирование ключевого слова голубя, J. Exp. Анальный. Behav. 11: BUZSAKI, G «Где это?» рефлекс: автоформирование ориентировочной реакции, Дж.Exp. Анальный. Behav. 37: ЭЛУЛ, Р. и МАРКИАФАВА, П. Л. Аккомодация глаза по отношению к поведению кошки. Архипелаг Курсив Biol. 102: ГРАСТЬЯН Э. и ВЕРЕЧКЕЙ Л. Эффекты пространственного разделения условного сигнала от подкрепления. Демонстрация условности ориентировочной реакции ориентировочного характера обусловленности. Behav. Biol. 10: HEARST, E Сигналы, условные направленные движения и типичные для вида предрасположенности к ответам у позвоночных, не являющихся млекопитающими.В П. Марлер и Х. С. Террас (ред.), Биология обучения, отчет об исследовании наук о жизни 29. Springer-Verlag, Berlin, p KONORSKI, J. Интегративная активность мозга. Университет Чикаго Пресс, Чикаго, 530 стр. 7. КОССУТ, М., МИХАЛЬСКИЙ, А. и ЗЕРНИЦКИЙ, Б. Глазной рефлекс следования у кошек, лишенных паттернов зрения с рождения. Brain Res. 141: КУПАЛОВ П.С., ЯРОСЛАВЦЕВА О.П. Возможность выработки условного ориентировочного рефлекса на функциональное состояние коры головного мозга.Труды Физиол. Лаборатория Павлова, Акад. Med. АН СССР 15: ЕАВИКА, З. Слуховые прицельные рефлексы: их определяющая роль в направленном инструментальном реагировании. Acta Neurobiol. Exp. 39: МАЖОРОВ Ф.П. История учения об условных рефлексах. Издат. АМН СССР, Москва, 374 с. 11. МИХАЛСИ А., КОССУТ М. и ЗЕРНИЦКИЙ Б. Глазной рефлекс следования, вызванный периферией сетчатки у кошки. Vision Res. 17: ПАВЛОВ И.П. Условные рефлексы. Исследование физиологической активности коры головного мозга.Oxford Univ. Press, Oxford, 430 p. 13. ПАВЛОВ И.П. Двадцатилетний опыт изучения высшей нервной деятельности (поведения) животных. Med. Лит., Ленинград, 236 с. 14. ROHRBAUGH, Дж. В. Ориентирующий рефлекс: производительность и проявления центральной нервной системы. В Р. Парасураман и Д. Р. Дэвис (ред.), Разновидности внимания. Акад. Press, Orlando, p SANTIBANEZ, G Прицельный рефлекс. Acta Neurobiol. Exp. 36: СИДДЛ, Д. А. Т., КУЙАК, М. и СТЕНФЕРТ КРОЗЕ, Б. Ориентирующий рефлекс. У А. Гейла и Дж.А. Эдвардс (редактор), физиологические корреляты человеческого поведения. Том 2. Акад. Press, Лондон, стр.

9 17. СОКОЛОВ Е. Н. Нейрональные модели и ориентировочный рефлекс. В М. А. Б. Бразье (ред.), Центральная нервная система и поведение. Фонд Джослии Мэйси-младшего, Нью-Йорк, СОКОЛОВ Е.Н. Восприятие и условный рефлекс. Pergamon Press, Oxford, 309 p. 19. СОКОЛОВ, Е.N Высшие нервные функции: ориентировочный рефлекс. Энн. Rev. Physiol. 25: СОКОЛОВ Е.Н. Нейрональные механизмы ориентировочного рефлекса. В работе Е. Н. Соколова и О. С. Виноградова (ред.), Нейронные механизмы ориентировочного рефлекса. Lawrence Erlbaum Associates, Publ. Hillsdale, New Jersey, p STgPIEN, I Магнитная реакция, симптом префронтальной абляции Acta Neurobiol. Exp. 34: YARBUS, A. L Движения глаз и зрение. Plenum Press, Нью-Йорк, 222 стр. 23. ЗЕРНИЦКИЙ, Б. Ориентирующая реакция гипернормальности у лобных кошек.Acta Neurobiol. Exp. 32: ЗЕРНИЦКИЙ, Б. Препарирование до тригемии. Архипелаг Курсив Biol. 124: ZERNICKI, B. и DREHER, B Исследования рефлекса зрительной фиксации. I. Общие свойства рефлекса ориентировочной фиксации у претригеминальных и интактных кошек. Acta Bid. Exp. 25: Принято 20 июля 1987 г.

,
Стереотипный ответ | биология | Британника
Стереотипный ответ , неизученная поведенческая реакция организма на некоторый раздражитель окружающей среды. Это адаптивный механизм, который может выражаться по-разному. Все живые организмы демонстрируют один или несколько типов стереотипной реакции.
Общие положения
Способность к необученному поведению определяется генетически во многом в том же смысле, что и положение, размер, форма и функция органов.Подобно структурным особенностям, стереотипные ответы являются результатом непрерывного процесса эволюционной модификации и уточнения. Те действия, которые наиболее успешно помогают животному или растению в его основных побуждениях (, например, размножение, поиск пищи, бегство от хищников), с наибольшей вероятностью будут сохранены в следующих поколениях. По мере изменения условий окружающей среды внутренне детерминированные реакции также изменяются, чтобы обеспечить продолжение вида.
Проблемы, возникающие при изучении стереотипных ответов, многочисленны и разнообразны.Особые реакции у животных не всегда поддаются идентификации в высокоразвитых формах, потому что усвоенные поведенческие паттерны скрывают лежащее в их основе необученное поведение; кроме того, стереотипные ответы служат строительными блоками инстинктивного поведения, сложность которого может затенять составные части (см. инстинкт). У низших животных, как и у растений, у которых обученное поведение отсутствует или отсутствует, анализ поведенческих механизмов ограничен тем фактом, что многие из наиболее фундаментальных клеточных процессов не совсем понятны.
Поведение животных как раздел психологии представляет собой слияние дисциплин этологии и сравнительной психологии. Большая часть новаторской работы по созданию стереотипных реакций животных была проделана этологами. В течение первой половины 20 века, когда были заложены многие основы экспериментальной психологии, этологи (по большей части европейцы) интересовались поведением насекомых, рыб и птиц и особенно интересовались эволюцией инстинктов. ,Психологами-сравнителями в этот период становления были в основном американцы. Они изучали в основном поведение обычных лабораторных животных, таких как морские свинки, мыши, крысы и обезьяны, и их интерес, как правило, был сосредоточен на влиянии окружающей среды на поведение, а не на генетическом влиянии. С 1950-х годов психологи в целом признали, что факторы окружающей среды и генетические факторы играют важную роль в любом биологическом явлении. Однако в результате раздельного развития этологии и сравнительной психологии возникли некоторые трудности в использовании терминологии.Немецко-американский биолог Жак Леб применил термин тропизм ко всем ориентированным движениям организмов и предположил, что все поведение состоит из тропизмов. Впоследствии, чтобы избежать путаницы, другие исследователи ввели термины «налоги» (в единственном числе: такси) и кинезы для обозначения реакций животных, не относящихся к оседлым, похожим на растения формам. Эти термины также применялись к определенным движениям растений. Хотя у растений происходит множество дискретных стереотипных ответных движений, особенно у высших форм, таких как цветковые растения, эти автономные движения обычно происходят слишком медленно, чтобы их можно было обнаружить путем случайного наблюдения.То, что движения растений или органов растений действительно имеют место, может быть наглядно продемонстрировано с помощью покадровой фотографии, при которой отдельные фотографии делаются через равные промежутки времени, такие как секунды или несколько дней или более. Затем фотографии сравниваются или просматриваются в быстрой последовательности как движущееся изображение.
Получите эксклюзивный доступ к контенту нашего 1768 First Edition с подпиской.
Подпишитесь сегодня
Типы стереотипных ответов
Стереотипный ответ у животных можно разделить на следующие четыре категории: неорганизованный или плохо организованный ответ, рефлекторные движения определенной части организма, рефлекторная активность всего организма и инстинкт.
Неорганизованные или плохо организованные реакции дают ранние эмбрионы или животные (например, губки), у которых отсутствует нервная система.
Собственно рефлексы или рефлекторно-дуговые движения включают такие реакции, как немедленное отдергивание руки при прикосновении к горячей поверхности. Основными компонентами рефлекторной дуги являются рецептор, или сенсорно-нервная клетка, которая воспринимает стимул, и аффектор, нервная клетка, которая непосредственно активирует мышцу. Это теоретический минимум, а не наблюдаемое функциональное расположение клеток в теле животного (см. Инстинкт: разновидности инстинктивного поведения).
Рефлексоподобная деятельность
Рефлекторная деятельность целых организмов может быть неориентированной или ориентированной. Неориентированные реакции включают кинезы — ненаправленное ускорение или замедление скорости передвижения или частоты перехода от состояния покоя к движению (ортокинез), или частоты или количества поворотов всего животного (клинокинез), скорость частоты зависит от интенсивности стимуляции. , Примеры ортокинеза наблюдаются у миног, которые более активны при высокой интенсивности света, и у тараканов, которые более активны при низкой интенсивности; плоские черви и многие виды личинок мух, среди других беспозвоночных, обладают ортокинезом.Клинокинез хорошо демонстрируют движения мокрицы ( Porcellio scaber ). Когда мокрицы помещаются в сухой воздух, они активно ползают, но без направления, пока постепенно не обезвоживаются. Когда мокрицы помещаются во влажный воздух, они сначала двигаются, но всякая активность, которую они проявляют, вскоре прекращается, и они затихают. Мыши, помещенные в контейнер с сухим воздухом на одном конце и влажным — на другом, постепенно собираются на влажном конце. Эта передача достигается скорее случайными, чем направленными движениями.
Ориентированная рефлекторная деятельность целых организмов включает тропизмы, налоги и ориентацию под углом. Тропизмы у животных — это направленные движения роста-искривления сидячих (, т. Е. сидячих) форм, которые приводят к одинаковой интенсивности стимуляции симметрично расположенных частей тела. Эти движения демонстрируют гидроидные животные, такие как Eudendrium .
.