8.5.8. Гомеостаз популяций. Общая экология
8.5.8. Гомеостаз популяций
Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит.
Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других – в смягченной, например в понижении плодовитости на основе условных рефлексов.
Рис. 129. Самоизреживание в древесных насаждениях (по Г. Ф. Морозову, 1928):
слева – господствующие и угнетенные деревья в ельнике; справа – ход изреживания стволов с возрастом у сосны (1), березы (2) и ели (3)
К жестким формам внутривидовой конкуренции следует отнести, например, явление самоизреживания у растений (рис. 129). При большой густоте всходов часть растений неминуемо погибает в результате угнетения физиологически более сильными соседями. Уменьшение числа растений происходит, даже если высеянные семена генетически однородны. В этом случае, по-видимому, имеют значение разница в размерах семян, во времени появления всходов, детали микроокружения. В одном из опытов с клевером Trifolium subterraneum через 84 дня после появления всходов на участке в 1 м2 из 1250 растений осталось 650, причем влияние вредителей было исключено.
У райграса многолетнего основной экологической единицей становится не особь, а побег. Обнаружено, что при разных нормах высева семян, от 6 до 180 кг/га, сначала густота побегов варьирует от 30 до 1070 на 100 см2, но затем во всех случаях становится равной примерно 500, т. е. в более редких посевах появляются новые побеги, а в более густых часть отмирает.
Регуляция плотности популяции у растений ввиду особенностей их роста происходит обычно не только путем изменения численности особей на единице площади, но и путем изменения вегетативной мощности каждого. В загущенных посевах растения менее облиственны, с меньшим количеством побегов. Общая их масса при увеличении плотности посевов сначала возрастает пропорционально количеству высеянных семян, а затем остается на постоянном уровне, тогда как средняя масса отдельных особей соответственно уменьшается. В данном случае стабилизируется не число особей в популяции, а общая листовая фотосинтезирующая поверхность растений.
У животных жесткие формы регуляции плотности популяций проявляются обычно лишь в тех случаях, когда запасы пищи, воды или других ресурсов резко ограничены, а животные либо не способны в данный период к поискам ресурсов на другой территории, либо эти поиски неэффективны. Например, в небольших пресноводных водоемах, где нет других видов рыб, популяции окуня могут поддерживать свое существование и регулировать плотность за счет питания взрослых собственной молодью. Мальки же растут за счет мелкого планктона, к питанию которым крупные окуни не приспособлены. Каннибализм – не частое явление в популяциях животных.
Особый интерес представляют некоторые, сравнительно редкие виды, у которых способность к умерщвлению конкурентов внутри популяции закрепляется эволюционно в их поведении и даже морфологии. Подобные примеры встречаются среди насекомых.
Для паразитоидных перепончатокрылых, откладывающих свои яйца в яйца или личинки других насекомых, запас пищи в хозяине весьма ограничен и дает возможность развиться чаще всего лишь одной особи паразита. У ряда видов личинки первого возраста имеют челюсти, ненужные при питании содержимым яйца, но приспособленные для схваток с конкурентами. Нападение личинок друг на друга обычно происходит до того, как они начинают кормиться. После уничтожения конкурента личинка линяет и ее острые челюсти сменяются мягкими и бугорковидными (рис. 130). У тех видов, самки которых способны распознать уже зараженных хозяев и не откладывать яйца в них повторно, личинкам не приходится конкурировать между собой, и они лишены острых челюстей.

Рис. 130. Личинки паразитоидных перепончатокрылых:
1– Opius fletcheri; 2 – Galesus sylvestrii (a – первый возраст, б – второй возраст)
Групповые паразиты, откладывающие по нескольку яиц в одно насекомое, нередко обладают способностью соразмерять число яиц с величиной жертвы. Однако при недостатке хозяев возможно перезаражение обнаруженных паразитами, при этом обеспеченность потомства кормом снижается. В таких случаях наблюдается уменьшение размеров личинок. Вышедшие из них имаго откладывают вдвое-втрое меньше яиц при сокращенных сроках жизни. Например, у самых мелких наездников-браконид происходит полная редукция яичников.
Плотность популяции у паразитических перепончатокрылых может регулироваться также через изменения в соотношении полов, что влияет на численность следующего поколения. У многих видов сильно различается смертность самцов и самок при перенаселении. Например, у некоторых браконид доля самок составляет около 70 % при слабом заражении насекомых-хозяев и падает ниже 10 % – при сильном. Кроме того, соотношение полов регулируется поведением самок. Для многих видов наездников установлено, что в мелких хозяевах, представляющих малый запас корма, самки откладывают преимущественно неоплодотворенные яйца, из которых у перепончатокрылых развиваются самцы. При учащении контактов самок друг с другом или при восприятии следовых запахов, оставляемых другими самками, среди откладываемых яиц также увеличивается доля неоплодотворенных.
Среди механизмов, задерживающих рост популяций, у многих видов большую роль играют химические взаимодействия особей. Так, вода аквариума, в котором содержались дафнии, способна задерживать рост представителей того же вида и сохраняет эту способность в течение нескольких дней. Головастики выделяют в воду частицы белковой природы, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких, так как устойчивость к одной и той же концентрации ингибитора находится в прямой зависимости от размеров. Один крупный головастик Rana pipiens может задержать рост всех других в 75-литровом аквариуме. Поколение, вышедшее в близкие сроки из икры, отложенной в одном водоеме, вскоре разделяется на две размерные группы: более крупные, продолжающие расти головастики и мелкие, затормозившие свой рост из-за неблагоприятной для них концентрации метаболита. Экологическая выгода такого разделения популяции в том, что особи с наследственно более быстрым темпом роста, используя в полной мере кормовые ресурсы водоема, получают возможность быстро завершить метаморфоз и в популяцию вливается полноценное пополнение. Оставшиеся мелкие головастики, после того как первая партия покинет водоем и в нем снизится концентрация ингибитора, также имеют шансы увеличиться в размерах и достичь стадии метаморфоза, но значительно позже. Эту вторую часть пополнения можно рассматривать как своего рода резерв, который вливается в популяцию лишь при достаточно благоприятных условиях (если не пересохнет временный водоем, если в нем продолжится воспроизводство водорослей – основной пищи головастиков и т. п.).
Выделение в окружающую среду продуктов, задерживающих рост, обнаружено у многих растений и водных животных, особенно у рыб.
Другой механизм ограничения численности популяций – такие изменения физиологии и поведения при увеличении плотности, которые в конечном счете приводят к проявлению инстинктов массовой миграции. В результате происходит выселение большей части популяции за пределы территории, занимаемой в оседлый период. Особенно ярко это проявляется у насекомых, которым свойственна фазовость – резкое изменение морфологии и физиологии особей в зависимости от плотности популяции (рис. 131). У перелетной саранчи-шистоцерки в постоянных очагах ее обитания в Индии, Пакистане, Восточной Африке и Аравии при низкой численности личинки одиночной фазы имеют ярко-зеленую, а взрослые – серовато-зеленую или бурую окраску. В годы массового размножения, которое наступает при благоприятном сочетании погодных условий, саранча переходит в стадную фазу. Личинки приобретают ярко-желтую, с черными пятнами окраску, взрослая неполовозрелая шистоцерка интенсивно розово-лиловая, половозрелая – лимонно-желтая. Изменяется и морфология особей: удлиняются надкрылья, меняются формы переднеспинки, киля, пропорции конечностей и т. п. Переход из одной фазы в другую занимает около трех поколений. Процесс стимулируется зрительным восприятием особой своего вида и контактами при помощи усиков. Это вызывает в организме насекомых целую серию реакций, приводящих к гормональным перестройкам, в которых участвуют эндокринные железы.

Рис. 131. Нимфы V возраста саранчи-шистоцерки (по Н. С. Щербиновскому, 1952): слева – стадная форма; справа – одиночная форма
Стадная фаза отличается повышенной возбудимостью и чрезвычайной прожорливостью. Плодовитость самок снижается, но они откладывают яйца с большим содержанием питательных веществ. Стадная саранча все время находится в состоянии миграционной активности. Личинки двигаются скоплениями – кулигами, а взрослые гигантскими стаями разлетаются на сотни и тысячи километров от мест постоянного обитания (рис. 132). Так, в конце прошлого столетия масса одной из стай шистоцерки, перелетевшей через Красное море, была определена не менее чем в 44 млн т.
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2–3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.

Рис. 132. Нашествие перелетной саранчи
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотности населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поселений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.
Территориальное поведение животных, выработавшееся в ходе эволюции как система инстинктов, – наиболее эффективный механизм сдерживания роста численности популяции на данной площади. Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводят к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства. Это же относится и к выведенному потомству, среди которого лишь некоторая часть вследствие естественной смертности взрослых занимает освобождающиеся участки.
Выселения как ответная реакция на растущую плотность популяции свойственны многим видам птиц и млекопитающих. Кроме обычной расселительной дисперсии молодняка, для ряда видов с резкими колебаниями численности характерны массовые перемещения – нашествия. Они возникают нерегулярно, лишь в годы вспышек размножения, и не имеют постоянного направления. Такие нашествия описаны, например, у тундровых леммингов, белок Сибири и Северной Америки и др. При нашествиях часть особей остается на месте, а среди эмигрантов преобладают молодые.
Повышение плотности популяции может сопровождаться такими изменениями в физиологии особей, которые ведут к снижению рождаемости и увеличению смертности. У млекопитающих известно явление стресса, которое впервые было описано в 1936 г. физиологом Г. Селье для человека. В ответ на отрицательное воздействие каких-либо факторов в организме возникают реакции двух типов: 1) специфические, зависящие от природы повреждающего агента (например, усиление теплопродукции при действии холода), и 2) неспецифическая реакция напряжения как общее усилие организма приспособиться к изменившимся условиям. Эта общая реакция складывается из ряда физиологических и морфологических изменений, которые постепенно развертываются как единый процесс. Реакция напряжения, или стресс, возникает в ответ на любые отрицательные воздействия среды, в том числе и на отклонение плотности популяции от оптимума.
Большую роль в развитии стресса играют сигналы коры головного мозга, меняющие активность гипоталамуса – центрального звена вегетативной нервной системы. В свою очередь, деятельность гипоталамуса вызывает изменения в функционировании гипофизарно-надпочечниковой гормональной системы. В состоянии стресса у животных сильно увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов, выделяемых этим органом, а также происходит целый ряд других изменений в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, резорбция эмбрионов, рано прекращается лактация, угасают инстинкты заботы о потомстве и т. п., уменьшается число выводков и количество в них молодых. В конечном счете все это приводит к торможению роста популяции. У мышевидных грызунов, содержащихся в клетках одинаковых размеров, четко проявляется обратно пропорциональная зависимость между численностью зверьков в клетке и массой их репродуктивных органов. В состоянии стресса у животных даже при достаточном снабжении кормом понижается устойчивость к вредным воздействиям среды, что увеличивает смертность.
От плотности населения зависит в первую очередь поведение животных. У многих видов в условиях скученности повышается уровень агрессивности, меняется реакция на особей противоположного пола, молодняк и т. д.
Стресс-реакция как механизм, регулирующий рождаемость, особенно отчетливо проявляется у животных с хорошо выраженной системой иерархического подчинения в группах.
Реакция напряжения свойственна подчиненным животным; у них также тормозится воспроизводительная функция. Доминирующие особи стресс-реакции не проявляют. В переуплотненных популяциях стресс распространяется на большую часть особей и, по-видимому, тормозит размножение.
Стрессовое состояние не вызывает необратимых изменений в половой системе, а приводит лишь к временному гормональному блокированию ее функции. После устранения перенаселенности способность к размножению может восстановиться в короткие сроки.
Закономерности стресса, вызываемого перенаселением, изучают в основном на лабораторных животных. Однако многочисленные факты, зарегистрированные в природных популяциях, позволяют предполагать, что в естественных условиях стресс играет немалую роль в регуляции численности и структуры популяций и поведения млекопитающих (рис. 133).

Рис. 133. Зависимость интенсивности размножения от плотности населения в популяции малого суслика (по М. Р. Магомедову, 1995)

Рис. 134. Динамика численности сибирского лемминга на Аляске (по Bunnel et all., 1975)
Например, в динамике популяций ряда тундровых леммингов зарегистрированы правильные циклы трех-четырехлетней периодичности с амплитудой колебания до 600 крат (рис. 134). Фаза пика в таких колебаниях ограничена обычно одним сезоном, затем следуют резкий спад, фаза депрессии численности и последующего нарастания. При нарастании численности регистрируется увеличение плодовитости, повышение скорости созревания молодняка, усложнение возрастной структуры популяции, снижение смертности новорожденных. В период пика отмечается резкое снижение воспроизводства и одновременно увеличивается смертность во всех возрастных группах. В яичниках самок всех возрастов обнаруживаются серьезные деструктивные изменения, наблюдается массовая гибель фолликулов на ранних стадиях развития. Год-два после спада численности общая интенсивность размножения остается средней, а смертность высокой, а затем вновь все показатели размножаемости начинают расти. У зверьков, родившихся при низкой численности популяции, восстанавливается нормальное функционирование яичников.
На разных этапах этого цикла сильно меняется гормональное состояние зверьков, относящихся к разным генерациям. В пик численности отмечается избыточная активность адреналовой и щитовидной желез, что резко тормозит воспроизводительные функции организма. В ходе цикла у сменяющихся поколений меняется не только функционирование отдельных желез, но и всей эндокринной системы животных. Через несколько поколений, в период минимума численности, состояние эндокринной системы нормализуется и обеспечивает восстановление эффективности процесса размножения. Конкретные причины таких гормональных различий у разных поколений связаны с тем, что жизнеспособность и эндокринные особенности организма формируются в эмбриональный период и во многом определяются физиологическим состоянием родителей.
Таким образом, динамику численности леммингов можно представить как авторегулируемый процесс, в котором большую роль играют эндокринные механизмы.
Все рассмотренные выше примеры взаимодействия между членами популяции, от «жестких» форм – прямого уничтожения одной особью другой – до снижения воспроизводительных способностей как условного рефлекса на повышение частоты контактов, представляют собой разные формы ограничения роста популяций. Эти тормозящие механизмы включаются до полного истощения ресурсов среды в ответ на сигналы, свидетельствующие об угрозе перенаселения.
Степень развития механизмов популяционного гомеостаза находится также в тесной связи с тем, насколько влияют на популяцию другие виды: конкуренты, хищники, паразиты. Общая регуляция численности популяций в природных сообществах происходит в результате сложных межвидовых и внутривидовых взаимоотношений.
Данный текст является ознакомительным фрагментом.
Читать книгу целиком
Поделитесь на страничке
Следующая глава >
32. Гомеостаз популяций. Механизмы гомеостаза и причины его нарушения.
Поддержание
определенной численности получило
название гомеостаза
популяций.
В
основе способностей популяций к
гомеостазу лежат изменения физиологических
особенностей, роста, поведения каждой
особи в ответ на увеличение или уменьшение
числа членов популяции, к которой она
принадлежит.
Механизмы
популяционного гомеостаза
зависят от экологической специфики
вида, его подвижности, степени воздействия
хищников и паразитов и др. У одних
видов они могут проявляться в жесткой
форме, приводя к гибели избытка особей,
у других — в смягченной, например в
понижении плодовитости на основе
условных рефлексов.
К
жестким формам внутривидовой, конкуренции
следует отнести, например, явление
самоизреживания у растений. При большой
густоте всходов часть растений неминуемо
погибает в результате угнетения
физиологически более сильными соседями.
Уменьшение числа растений происходит,
даже если высеянные семена генетически
однородны. В этом случае, по-видимому,
имеют значение разница в размерах,
во времени появления всходов, детали
микроокружения. В одном из опытов с
клевером Trifoliurn
sim-terraneum
через 84 дня после появления всходов на
участке в 1 м2
из
1250 растений осталось 650, причем влияние
вредителей было исключено
У
райграса многолетнего основной
экологической единицей становится не
особь, а побег. Обнаружено, что при разных
нормах высева семян, от 6 до 180 кг/га,
сначала густота побегов варьирует от
30 до 1070 на 100 см2,
но затем во всех случаях становится
равной примерно 500, т. е. в более редких
посевах появляются новые побеги, а
в более густых часть отмирает.
Регуляция
плотности популяции у растений ввиду
особенностей их роста происходит
обычно не только путем изменения
численности особей на единице площади,
но и путем изменения вегетативной
мощности каждого. В загущенных посевах
растения менее облиственны, с меньшим
количеством побегов. Общая их масса
при увеличении плотности посевов сначала
возрастает пропорционально количеству
высеянных семян, а затем остается на
постоянном уровне, тогда как средняя
масса отдельных особей соответственно
уменьшается. В данном случае стабилизируется
не число особей в популяции, а общая
листовая фотосинтезкрующая поверхность
растений.
У
животных жесткие формы регуляции
плотности популяций проявляются обычно
лишь в тех случаях, когда запасы пищи,
воды или других ресурсов резко
ограничены, а животные либо не способны
в данный период к поискам ресурсов на
другой территории, либо эти поиски
неэффективны. Например, в небольших
пресноводных водоемах, где нет других
видов рыб, популяции окуня могут
поддерживать свое существование и
регулировать плотность за счет питания
взрослых собственной молодью. Мальки
же растут за счет мелкого планктона, к
питанию которым крупные окуни не
приспособлены. Каннибализм возникает
иногда в период длительного голодания
и в популяциях хищных млекопитающих
Особый
интерес представляют некоторые,
сравнительно редкие виды, у которых
способность к умерщвлению конкурентов
внутри популяции закрепляется эволюционно
в их поведении и даже морфологии. Подобные
примеры встречаются среди насекомые.
Для паразитоидных перепончатокрылых,
откладывающих свои яйца в яйца или
личинки других насекомых, запас пищи в
хозяине весьма ограничен и дает
возможность развиться чаще всего лишь
одной особи паразита. У ряда видов
личинки первого возраста имеют
челюсти, ненужные при питании содержимым
яйца, но приспособленные для схваток с
конкурентами. Нападение личинок друг
на друга обычно происходит до того, как
они начинают кормиться. После уничтожения
конкурента личинка линяет и ее острые
челюсти сменяются мягкими и бугорковидными.
У тех видов, самки которых способны
распознать уже зараженных хозяев и не
откладывать яйца в них повторно, личинкам
не приходится конкурировать между
собой, и они лишены острых челюстей.
Групповые
паразиты, откладывающие по нескольку
яиц в одно насекомое, нередко обладают
способностью соразмерять число яиц
с величиной жертвы. Однако при недостатке
хозяев возможно перезаражение
обнаруженных паразитами, при этом
обеспеченность потомства кормом
снижается. В таких случаях наблюдается
уменьшение размеров личинок. Вышедшие
из них имаго откладывают вдвое-втрое
меньше яиц при сокращенных сроках жизни.
Например, у самых мелких наездииков-браконид
происходит полная редукция яичников.
Плотность
популяции у паразитических перепончатокрылых
может регулироваться также через
изменения в соотношении полов, что
влияет на численность следующего
поколения. У многих видов сильно
различается смертность самцов и самок
при перенаселении. Например, у
некоторых браконид доля самок составляет
около 70% при слабом заражении
насекомых-хозяев и падает ниже 10%—при
сильном. Кроме того, соотношение полов
регулируется поведением самок. Для
многих видов наездников установлено,
что в мелких хозяевах, представляющих
малый запас корма, самки откладывают
преимущественно неоплодотворенные
яйца, из которых у перепончатокрылых
развиваются самцы. При учащении контактов
самок друг с другом или при восприятии
следовых запахов, оставляемых другими
самками, среди откладываемых яиц
также увеличивается доля неоплодотворенных.
Среди
механизмов, задерживающих рост популяций,
у многих
видов
большую роль играют химические
взаимодействия особей. Так, вода
аквариума, в котором содержались дафнии,
способна задерживать рост представителей
того же вида и сохраняет эту способность
в течение нескольких дней. Головастики
выделяют в воду частицы белковой природы,
которые задерживают рост других
головастиков. Чем крупнее особь, тем
сильнее она воздействует на более
мелких, так как устойчивость к одной и
той же концентрации ингибитора находится
в прямой зависимости от размеров
Один крупный головастик Rana
pipiens
может задержать рост всех других в
75-литровом аквариуме. Поколение, вышедшее
в близкие сроки из икры, отложенной в
одном водоеме, вскоре разделяется на
две размерные группы: более крупные,
продолжающие расти головастики и
мелкие, затормозившие свой рост из-за
неблагоприятной для них концентрации
метаболита. Экологическая выгода
такого разделения популяции в том, что
особи с наследственно более быстрым
темпом роста, используя в полной мере
кормовые ресурсы водоема, получают
возможность быстро завершить
метаморфоз и в популяцию вливается
полноценное пополнение. Оставшиеся
мелкие головастики, после того как
первая партия покинет водоем и в нем
снизится концентрация ингибитора, также
имеют шансы увеличиться в размерах и
достичь стадии метаморфоза, но
значительно позже. Эту вторую часть
пополнения можно рассматривать как
своего рода резерв, который вливается
в популяцию лишь при достаточно
благоприятных условиях (если не
пересохнет временный водоем, если о
нем продолжится воспроизводство
водорослей —основной пищи головастиков
и т. п.).
Выделение в
окружающую среду продуктов, задерживающих
рост, обнаружено у многих растений и
водных животных, особенно у рыб.
Другой
механизм ограничения численности
популяций — такие изменения физиологии
и поведения при увеличении плотности,
которые в конечном счете приводят к
проявлению инстинктов массовой
миграции. В результате происходит
выселение большей части популяции за
пределы территории, занимаемой в оседлый
период. Особенно ярко это проявляется
у насекомых, которым свойственна
фазовость
—
резко разнокачественное состояние
особей в зависимости от плотности
популяции. У перелетной саранчи-шисгоцерки
в постоянных очагах ее обитания в Индии,
Пакистане, Восточной Африке и Аравии
при низкой численности личинки одиночной
фазы имеют ярко-зеленую, а взрослые —
серовато-зеленую или бурую окраску. В
годы массового размножения, которое
наступает при благоприятном сочетании
погодных условий, саранча переходит в
стадную фазу. Личинки приобретают
ярко-желтую, с черными пятнами окраску,
взрослая неполовозрелая шистоцерка
интенсивно розово-лиловая, половозрелая
— лимонно-желтая. Изменяется и морфология
особей: удлиняются надкрылья, меняются
формы переднеспинки, киля, пропорции
конечностей и т. п. Переход из одной фазы
в другую занимает около трех поколений.
Процесс стимулируется зрительным
восприятием особей своего вида и
контактами при помощи усиков. Это
вызывает в организме насекомых целую
серию реакций, приводящих к гормональным
перестройкам, в которых участвуют
эндокринцые железы.
Стадная
фаза отличается повышенной возбудимостью
и чрезвычайной прожорливостью.
Плодовитость самок снижается, но
они
откладывают яйца с большим содержанием
питательных веществ. Стадная саранча
все время находится в состоянии
миграционной активности. Личинки
двигаются скоплениями — кулигами,
а взрослые гигантскими стаями разлетаются
на сотни и тысячи километров от мест
постоянного обитания. Так, в конце
прошлого столетия масса одной из стай
шнстоцерки, перелетевшей через Красное
море, была определена не менее чем в 44
млн. т.
На
границах своего временного ареала
перелетная саранча не может размножаться,
и эти очаги вскоре затухают. Стаи или
погибают, или, постепенно разреживаясь,
начинают откочевку к зоне постоянных
очагов, В изреженных популяциях вновь
происходит переход к одиночной фазе,
через 2—3 промежуточных поколения.
Таким образом, разлет стай саранчи не
обеспечивает формирования новых
постоянных популяций, а практически
служит лишь механизмом снятия
перенаселенности в местах, благоприятных
для размножения. При этом гибнет
гигантское количество насекомых.
Стаи, несущие на своем пути неисчислимые
бедствия, сами оказываются обреченными.
Явление
фазовости обнаружено не только у стадных
саранчовых, но и у других беспозвоночных.
У тлей увеличение плотности населения
вызывает появление крылатой фазы и
разлет насекомых с образованием
новых поколений. Обычно тли дают
несколько поколений бескрылых самок,
но в условиях постоянного перенаселения
крылатые самки развиваются в каждом
поколении. У ряда амеб химические
изменения состава водной среды, вызванные
переуплотнением популяции, стимулируют
переход в подвижную жгутиковую
стадию. В результате происходит быстрое
рассредоточение особей в пространстве.
Территориальное
поведение животных, выработавшееся в
ходе эволюции как система инстинктов,—-
наиболее эффективный механизм
сдерживания роста численности популяции
на данной площади. Мечение и охрана
участков, не допускающие размножения
на них «чужих» особей, приводят к
рациональному использованию территории.
Избыточная часть популяции при этом не
размножается или вынуждена выселяться
за пределы занятого пространства.
Это же относится и к выведенному
потомству, среди которого лишь некоторая
часть вследствие естественной смертности
занимает освобождающиеся участки.
Выселения
как ответная реакция на растущую
плотность популяции свойственны
многим видам птиц и млекопитающих.
Кроме обычной расселительной дисперсии
молодняка, для ряда видов с резкими
Колебаниями численности характерны
массовые перемещения— нашествия.
Они
возникают нерегулярно, лишь в годы
вспышек размножения, и не имеют постоянного
направления. Такие нашествия описаны,
например, у тундровых леммингов, белок
Сибири и Северной Америки и др. При
нашествиях часть особей остается на
месте, а среди эмигрантов преобладают
молодые.
Повышение
плотности популяции может сопровождаться
такими изменениями в физиологии
особей, которые ведут к снижению
рождаемости и увеличению смертности.
У млекопитающих известно явление
стресса,
которое
впервые было описано в 1936г. физиологом
Г. Селье для человека В ответ на
отрицательное воздействие каких-либо
факторов в организме возникают реакции
двух типов:
1)
специфические, зависящие от природы
повреждающего агента (например,
усиление теплопродукции при действии
холода), и
2)
неспецифическая реакция напряжения
как общее усилие организма приспособиться
к изменившимся условиям. Эта общая
реакция складывается из ряда физиологических
и морфологических изменений, которые
постепенно развертываются как единый
процесс. Реакция напряжения, или стресс,
возникает в ответ на любые отрицательные
воздействия среды, в том числе и на
отклонение плотности популяции от
оптимума.
Большую
роль в развитии стресса играют сигналы
коры головного мозга, меняющие
активность гипоталамуса — центрального
звена вегетативной нервной системы. В
свою очередь, деятельность гипоталамуса
вызывает изменения в функционировании
гипофизарно-надпочечниковой гормональной
системы. В состоянии стресса у животных
сильно увеличивается кора надпочечников
и повышается концентрация кортикостероидных
гормонов, выделяемых этим органом,
а также происходит целый ряд других
изменений в гормональном равновесии
организма. У самок в популяции
учащаются нарушения овуляции, резорбция
эмбрионов, рано прекращается лактация,
угасают инстинкты заботы о потомстве
и т. п., уменьшается число выводков и
количество в них молодых. В конечном
счете все это приводит к торможению
роста популяции. У мышевидных грызунов,
содержащихся в клетках одинаковых
размеров, четко проявляется обратно
пропорциональная зависимость между
численностью зверьков в клетке и массой
их репродуктивных органов. В состоянии
стресса у животных даже при достаточном
снабжении кормом понижается устойчивость
к вредным воздействиям среды, что
увеличивает смертность.
От
плотности населения зависит в первую
очередь поведение животных. У многих
видов в условиях скученности повышается
уровень агрессивности, меняется реакция
на особей противоположного пола,
молодняк и т. д. Нарушение сложившихся
стереотипов поведения приводит
большинство особей к состоянию стресса.
Стресс-реакция
как механизм, регулирующий рождаемость,
особенно отчетливо проявляется у
животных с хорошо выраженной системой
иерархического подчинения в группах.
Реакция
напряжения свойственна подчиненным
животным; у них также тормозится
воспроизводительная функция. Доминирующие
особи стресс-реакции не проявляют. В
переуплотненных популяциях стресс
распространяется на большую часть
особей и, по-видимому, тормозит размножение.
Стрессовое
состояние не вызывает необратимых
изменений в половой системе, а приводит
лишь к временному гормональному
блокированию ее функции. После
устранения перенаселенности способность
к размножению может восстановиться в
короткие сроки
Закономерности
стресса, вызываемого перенаселением,
изучают в основном на лабораторных
животных. Однако многочисленные
факты, зарегистрированные в природных
популяциях, позволяют предполагать,
что в естественных условиях стресс
играет немалую роль в регуляции
численности и структуры популяций и
поведения млекопитающих.
Например,
в динамике популяций ряда тундровых
леммингов зарегистрированы правильные
циклы трех-четырехлетней периодичности
с амплитудой колебания до 600 крат. Фаза
пика в каких колебаниях ограничена
обычно одним сезоном, затем следуют
резкий спад, фаза депрессии численности
и последующего нарастания При
нарастании численности регистрируется
увеличение плодовитости, повышение
скорости созревания молодняка, усложнение
возрастной структуры популяции, снижение
смертности новорожденных. В период
пика отмечается резкое снижение
воспроизводства и одновременно
увеличивается смертность во всех
возрастных группах. В яичниках самок
всех возрастов обнаруживаются
серьезные деструктивные изменения,
наблюдается массовая гибель фолликулов
на ранних стадиях развития. Год-два
после спада численности общая интенсивность
размножения остается средней, а
смертность высокой, а затем вновь все
показателя размножаемости начинают
расти. У зверьков, родившихся при низкой
численности популяции, восстанавливается
нормальное функционирование яичников.
На
разных этапах этого цикла сильно меняется
гормональное состояние зверьков,
относящихся к разным генерациям. В пик
численности отмечается избыточная
активность адреналовой и щитовидной
желез, что резко тормозит воспроизводительные
функции организма. В ходе цикла у
сменяющихся поколений меняется не
только функционирование отдельных
желез, но и всей эндокринной системы
животных. Через несколько поколений, в
период минимума численности, состояние
эндокринной системы нормализуется и
обеспечивает восстановление эффективности
процесса размножения. Конкретные
причины таких гормональных различий у
разных поколений связаны с тем, что
жизнеспособность и эндокринные
особенности организма формируются -в
эмбриональной
период и во многом определяются
физиологическим состоянием родителей.
Таким образом,
регуляцию численности леммингов можно
представить как авторегулируемый
процесс, в котором большую роль играют
эндокринные механизмы.
Все
рассмотренные выше примеры взаимодействия
между членами популяции, от «жестких»
форм — прямого уничтожения одной
особью другой — до снижения
воспроизводительных способностей
как условного рефлекса на повышение
частоты контактов, представляют собой
разные формы ограничения роста популяций.
Эти тормозящие механизмы включаются
до полного истощения ресурсов среды в
ответ на сигналы, свидетельствующие об
угрозе перенаселения.
Степень развития
механизмов популяционного гомеостаза
находится также в тесной связи с тем,
насколько влияют на популяцию другие
виды: конкуренты, хищники, паразиты
Общая регуляция численности популяций
в природных сообществах происходит
в результате сложных межвидовых и
внутривидовых взаимоотношений.
Гомеостаз популяции — Студопедия
Популяции организмов обладают способностью к естественному регулированию численности (плотности). Плотность популяции при более или менее значительных колебаниях остается в устойчивом состоянии между своим верхним и нижним пределами, что обеспечивается действием определенных приспособительных механизмов.
Тенденция популяций поддерживать внутреннюю стабильность с помощью собственных регулирующих механизмов называется гомеостазом, а колебания численности популяций в пределах какой-то средней величины -их динамическим равновесием. Все биологические системы характеризуются способностью к гомеостазу, т.е. к саморегуляции. С помощью саморегуляции поддерживается в целом существование каждой системы — ее состав и структура, внутренние связи и преобразования в пространстве и во времени.
Гомеостатическими являются, прежде всего, отдельные особи, а затем популяции. Саморегулирующиеся системы не замкнуты, они активно взаимодействуют с внешней средой и, следовательно, подвержены изменениям. Саморегуляция — необходимое приспособление организмов для поддержания жизни в постоянно меняющихся условиях. Саморегулирование популяции осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами. С одной стороны, это биотический потенциал, составляющий совокупность всех факторов, способствующих увеличению численности популяции, а с другой — это сопротивление среды — совокупность факторов, снижающих численность популяции (рис. 3.6).
Изменение численности зависит от целого ряда факторов среды – абиотических, биотических и антропогенных. Однако всегда можно выделить ключевой фактор, наиболее сильно влияющий на рождаемость, смертность, миграцию особей и т.д. Это так называемые регулирующие факторы. Они работают по принципу обратной отрицательной связи: чем значительнее численность, тем сильнее срабатывают механизмы, обуславливающие ее снижение, и наоборот, – при низкой численности сила этих механизмов ослабевает, и создаются условия для более полной реализации биотического потенциала.
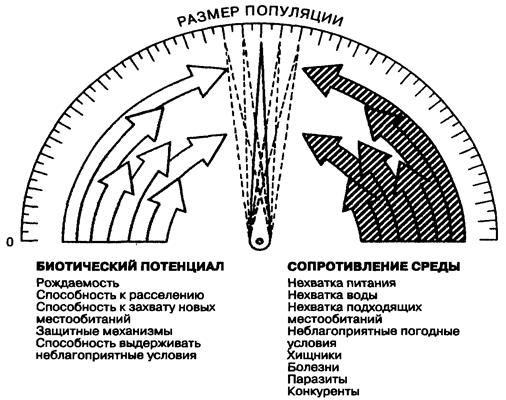
Рис. 3.6. Равновесное состояние популяции, определяемое
соотношением факторов, увеличивающих или снижающих
ее численность
Факторы, регулирующие плотность популяции, делятся на зависимые и независимые от плотности. Зависимые от плотности факторы изменяются вместе с изменением плотности, к ним относятся биотические факторы. Независимые от плотности факторыостаются постоянными с изменением плотности, это абиотические факторы.
Популяции многих видов организмов способны к саморегуляции своей численности, при которой на изменении численности популяции сказывается изменение качества особей.
Выделяют три механизма торможения роста численности популяций:
1) при возрастании плотности повышается частота контактов между особями, что вызывает у них стрессовое состояние, уменьшающее рождаемость и повышающее смертность;
2) при возрастании плотности усиливается эмиграция в новые местообитания, краевые зоны, где условия менее благоприятны, и смертность увеличивается;
3) при возрастании плотности происходят изменения генетического состава популяции, например, быстро размножающиеся особи заменяются медленно размножающимися.
Примером регулирующих факторов являются взаимоотношения организмов типа «хищник – жертва», «паразит – хозяин» и конкуренция. Высокая численность жертвы создает условия (пищевые) для размножения хищника. Последний, в свою очередь, увеличив численность, снижает количество жертвы. Численность обоих видов в результате этого носит синхронно-колебательный характер.
Действие регулирующих факторов можно рассматривать на уровне межвидовых и внутривидовых (внутрипопуляционных) взаимоотношений организмов. К межвидовым механизмам гомеостаза относятся взаимоотношения хищник — жертва, паразит — хозяин и конкуренция. Конкуренция лежит и в основе внутрипопуляционного гомеостаза. Она может проявляться в жестких и смягчающих формах. Жесткие формы заканчиваются гибелью части особей.
Например, в растительном мире внутривидовая конкуренция проявляется в самоизреживании фитоценозов. Па стадии всходов и молодых растений в лесных сообществах на 1 га насчитывается до нескольких сотен тысяч древесных растений. К возрасту спелости (100 — 120 лет для хвойных видов и 50 — 70 лет для лиственных) число экземпляров обычно не превышает 1000 на 1 га, но чаще — несколько сотен. Остальные погибают в результате острой конкуренции.
В животном мире результат острой внутривидовой борьбы проявляется в форме каннибализма (поедания себе подобных). Например, явления каннибализма характерны для некоторых грызунов, личинок насекомых (майского жука), хищных рыб — щук, окуней, корюшки, трески, наваги и других.
Смягченные формы внутривидовой конкуренции проявляются обычно через ослабление части особей, выключение их из процессов размножения.
Регуляция поддерживает численность популяции в пределах определенных границ, а при нарушении этих границ приводит к тому, что численность стремится вернуться в эти пределы. Это свидетельствует о важнейшей роли популяции как в генетико-эволюционном смысле, так и в чисто экологическом, как элементарной единицы эволюционного процесса, и об исключительной важности событий, протекающих на этом уровне биологической организации. Понимание механизмов регуляции численности популяций чрезвычайно важно для возможности управления этими процессами.
Деятельность человека часто сопровождается сокращением численности популяций многих видов. Причины этого в чрезмерном истреблении особей, ухудшении условий жизни вследствие загрязнения окружающей среды, беспокойства животных, особенно в период размножения, сокращение ареала и т.д. В природе нет и не может быть «хороших» и «плохих» видов, все они необходимы для ее нормального развития. В настоящее время остро стоит вопрос сохранения биологического разнообразия. Сокращение генофонда живой природы может привести к трагическим последствиям. Международный союз охраны природы и природных ресурсов (МСОП) издает «Красную книгу», где регистрирует следующие виды: исчезающие, редкие, сокращающиеся, неопределенные и «черный список» безвозвратно исчезнувших видов.
В целях сохранения видов человек использует различные способы регулирования численности популяции: правильное ведение охотничьего хозяйства и промыслов (установление сроков и угодий охоты и отлова рыбы), запрещение охоты на некоторые виды животных, регулирование вырубки леса и др. В то же время деятельность человека создает условия для появления новых форм организмов или развития старых видов, к сожалению, часто вредных для человека: болезнетворных микроорганизмов, вредителей сельскохозяйственных культур и т.д.
Таким образом, поскольку в настоящее время нарушения гомеостатических механизмов вызываются в большинстве случаев антропогенными факторами, то одной из важнейших задач человека является исключение или резкое снижение действия подобных факторов.
Гомеостаз популяций — Студопедия
Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит.
Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других – в смягченной, например в понижении плодовитости на основе условных рефлексов.
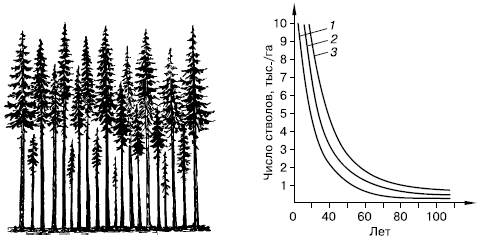
Рис. 129. Самоизреживание в древесных насаждениях (по Г. Ф. Морозову, 1928):
слева – господствующие и угнетенные деревья в ельнике; справа – ход изреживания стволов с возрастом у сосны (1), березы (2) и ели (3)
К жестким формам внутривидовой конкуренции следует отнести, например, явление самоизреживания у растений (рис. 129). При большой густоте всходов часть растений неминуемо погибает в результате угнетения физиологически более сильными соседями. Уменьшение числа растений происходит, даже если высеянные семена генетически однородны. В этом случае, по-видимому, имеют значение разница в размерах семян, во времени появления всходов, детали микроокружения. В одном из опытов с клевером Trifolium subterraneum через 84 дня после появления всходов на участке в 1 м2 из 1250 растений осталось 650, причем влияние вредителей было исключено.
У райграса многолетнего основной экологической единицей становится не особь, а побег. Обнаружено, что при разных нормах высева семян, от 6 до 180 кг/га, сначала густота побегов варьирует от 30 до 1070 на 100 см2, но затем во всех случаях становится равной примерно 500, т. е. в более редких посевах появляются новые побеги, а в более густых часть отмирает.
Регуляция плотности популяции у растений ввиду особенностей их роста происходит обычно не только путем изменения численности особей на единице площади, но и путем изменения вегетативной мощности каждого. В загущенных посевах растения менее облиственны, с меньшим количеством побегов. Общая их масса при увеличении плотности посевов сначала возрастает пропорционально количеству высеянных семян, а затем остается на постоянном уровне, тогда как средняя масса отдельных особей соответственно уменьшается. В данном случае стабилизируется не число особей в популяции, а общая листовая фотосинтезирующая поверхность растений.
У животных жесткие формы регуляции плотности популяций проявляются обычно лишь в тех случаях, когда запасы пищи, воды или других ресурсов резко ограничены, а животные либо не способны в данный период к поискам ресурсов на другой территории, либо эти поиски неэффективны. Например, в небольших пресноводных водоемах, где нет других видов рыб, популяции окуня могут поддерживать свое существование и регулировать плотность за счет питания взрослых собственной молодью. Мальки же растут за счет мелкого планктона, к питанию которым крупные окуни не приспособлены. Каннибализм – не частое явление в популяциях животных.
Особый интерес представляют некоторые, сравнительно редкие виды, у которых способность к умерщвлению конкурентов внутри популяции закрепляется эволюционно в их поведении и даже морфологии. Подобные примеры встречаются среди насекомых.
Для паразитоидных перепончатокрылых, откладывающих свои яйца в яйца или личинки других насекомых, запас пищи в хозяине весьма ограничен и дает возможность развиться чаще всего лишь одной особи паразита. У ряда видов личинки первого возраста имеют челюсти, ненужные при питании содержимым яйца, но приспособленные для схваток с конкурентами. Нападение личинок друг на друга обычно происходит до того, как они начинают кормиться. После уничтожения конкурента личинка линяет и ее острые челюсти сменяются мягкими и бугорковидными (рис. 130). У тех видов, самки которых способны распознать уже зараженных хозяев и не откладывать яйца в них повторно, личинкам не приходится конкурировать между собой, и они лишены острых челюстей.
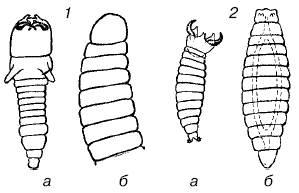
Рис. 130. Личинки паразитоидных перепончатокрылых:
1– Opius fletcheri; 2 – Galesus sylvestrii (a – первый возраст, б – второй возраст)
Групповые паразиты, откладывающие по нескольку яиц в одно насекомое, нередко обладают способностью соразмерять число яиц с величиной жертвы. Однако при недостатке хозяев возможно перезаражение обнаруженных паразитами, при этом обеспеченность потомства кормом снижается. В таких случаях наблюдается уменьшение размеров личинок. Вышедшие из них имаго откладывают вдвое-втрое меньше яиц при сокращенных сроках жизни. Например, у самых мелких наездников-браконид происходит полная редукция яичников.
Плотность популяции у паразитических перепончатокрылых может регулироваться также через изменения в соотношении полов, что влияет на численность следующего поколения. У многих видов сильно различается смертность самцов и самок при перенаселении. Например, у некоторых браконид доля самок составляет около 70 % при слабом заражении насекомых-хозяев и падает ниже 10 % – при сильном. Кроме того, соотношение полов регулируется поведением самок. Для многих видов наездников установлено, что в мелких хозяевах, представляющих малый запас корма, самки откладывают преимущественно неоплодотворенные яйца, из которых у перепончатокрылых развиваются самцы. При учащении контактов самок друг с другом или при восприятии следовых запахов, оставляемых другими самками, среди откладываемых яиц также увеличивается доля неоплодотворенных.
Среди механизмов, задерживающих рост популяций, у многих видов большую роль играют химические взаимодействия особей. Так, вода аквариума, в котором содержались дафнии, способна задерживать рост представителей того же вида и сохраняет эту способность в течение нескольких дней. Головастики выделяют в воду частицы белковой природы, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких, так как устойчивость к одной и той же концентрации ингибитора находится в прямой зависимости от размеров. Один крупный головастик Rana pipiens может задержать рост всех других в 75-литровом аквариуме. Поколение, вышедшее в близкие сроки из икры, отложенной в одном водоеме, вскоре разделяется на две размерные группы: более крупные, продолжающие расти головастики и мелкие, затормозившие свой рост из-за неблагоприятной для них концентрации метаболита. Экологическая выгода такого разделения популяции в том, что особи с наследственно более быстрым темпом роста, используя в полной мере кормовые ресурсы водоема, получают возможность быстро завершить метаморфоз и в популяцию вливается полноценное пополнение. Оставшиеся мелкие головастики, после того как первая партия покинет водоем и в нем снизится концентрация ингибитора, также имеют шансы увеличиться в размерах и достичь стадии метаморфоза, но значительно позже. Эту вторую часть пополнения можно рассматривать как своего рода резерв, который вливается в популяцию лишь при достаточно благоприятных условиях (если не пересохнет временный водоем, если в нем продолжится воспроизводство водорослей – основной пищи головастиков и т. п.).
Выделение в окружающую среду продуктов, задерживающих рост, обнаружено у многих растений и водных животных, особенно у рыб.
Другой механизм ограничения численности популяций – такие изменения физиологии и поведения при увеличении плотности, которые в конечном счете приводят к проявлению инстинктов массовой миграции. В результате происходит выселение большей части популяции за пределы территории, занимаемой в оседлый период. Особенно ярко это проявляется у насекомых, которым свойственна фазовость – резкое изменение морфологии и физиологии особей в зависимости от плотности популяции (рис. 131). У перелетной саранчи-шистоцерки в постоянных очагах ее обитания в Индии, Пакистане, Восточной Африке и Аравии при низкой численности личинки одиночной фазы имеют ярко-зеленую, а взрослые – серовато-зеленую или бурую окраску. В годы массового размножения, которое наступает при благоприятном сочетании погодных условий, саранча переходит в стадную фазу. Личинки приобретают ярко-желтую, с черными пятнами окраску, взрослая неполовозрелая шистоцерка интенсивно розово-лиловая, половозрелая – лимонно-желтая. Изменяется и морфология особей: удлиняются надкрылья, меняются формы переднеспинки, киля, пропорции конечностей и т. п. Переход из одной фазы в другую занимает около трех поколений. Процесс стимулируется зрительным восприятием особой своего вида и контактами при помощи усиков. Это вызывает в организме насекомых целую серию реакций, приводящих к гормональным перестройкам, в которых участвуют эндокринные железы.
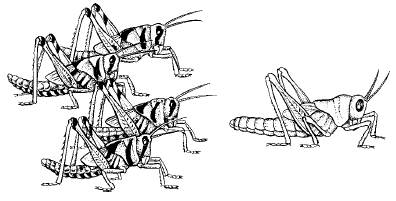
Рис. 131. Нимфы V возраста саранчи-шистоцерки (по Н. С. Щербиновскому, 1952): слева – стадная форма; справа – одиночная форма
Стадная фаза отличается повышенной возбудимостью и чрезвычайной прожорливостью. Плодовитость самок снижается, но они откладывают яйца с большим содержанием питательных веществ. Стадная саранча все время находится в состоянии миграционной активности. Личинки двигаются скоплениями – кулигами, а взрослые гигантскими стаями разлетаются на сотни и тысячи километров от мест постоянного обитания (рис. 132). Так, в конце прошлого столетия масса одной из стай шистоцерки, перелетевшей через Красное море, была определена не менее чем в 44 млн т.
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2–3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.

Рис. 132. Нашествие перелетной саранчи
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотности населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поселений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.
Территориальное поведение животных, выработавшееся в ходе эволюции как система инстинктов, – наиболее эффективный механизм сдерживания роста численности популяции на данной площади. Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводят к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства. Это же относится и к выведенному потомству, среди которого лишь некоторая часть вследствие естественной смертности взрослых занимает освобождающиеся участки.
Выселения как ответная реакция на растущую плотность популяции свойственны многим видам птиц и млекопитающих. Кроме обычной расселительной дисперсии молодняка, для ряда видов с резкими колебаниями численности характерны массовые перемещения – нашествия. Они возникают нерегулярно, лишь в годы вспышек размножения, и не имеют постоянного направления. Такие нашествия описаны, например, у тундровых леммингов, белок Сибири и Северной Америки и др. При нашествиях часть особей остается на месте, а среди эмигрантов преобладают молодые.
Повышение плотности популяции может сопровождаться такими изменениями в физиологии особей, которые ведут к снижению рождаемости и увеличению смертности. У млекопитающих известно явление стресса, которое впервые было описано в 1936 г. физиологом Г. Селье для человека. В ответ на отрицательное воздействие каких-либо факторов в организме возникают реакции двух типов: 1) специфические, зависящие от природы повреждающего агента (например, усиление теплопродукции при действии холода), и 2) неспецифическая реакция напряжения как общее усилие организма приспособиться к изменившимся условиям. Эта общая реакция складывается из ряда физиологических и морфологических изменений, которые постепенно развертываются как единый процесс. Реакция напряжения, или стресс, возникает в ответ на любые отрицательные воздействия среды, в том числе и на отклонение плотности популяции от оптимума.
Большую роль в развитии стресса играют сигналы коры головного мозга, меняющие активность гипоталамуса – центрального звена вегетативной нервной системы. В свою очередь, деятельность гипоталамуса вызывает изменения в функционировании гипофизарно-надпочечниковой гормональной системы. В состоянии стресса у животных сильно увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов, выделяемых этим органом, а также происходит целый ряд других изменений в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, резорбция эмбрионов, рано прекращается лактация, угасают инстинкты заботы о потомстве и т. п., уменьшается число выводков и количество в них молодых. В конечном счете все это приводит к торможению роста популяции. У мышевидных грызунов, содержащихся в клетках одинаковых размеров, четко проявляется обратно пропорциональная зависимость между численностью зверьков в клетке и массой их репродуктивных органов. В состоянии стресса у животных даже при достаточном снабжении кормом понижается устойчивость к вредным воздействиям среды, что увеличивает смертность.
От плотности населения зависит в первую очередь поведение животных. У многих видов в условиях скученности повышается уровень агрессивности, меняется реакция на особей противоположного пола, молодняк и т. д.
Стресс-реакция как механизм, регулирующий рождаемость, особенно отчетливо проявляется у животных с хорошо выраженной системой иерархического подчинения в группах.
Реакция напряжения свойственна подчиненным животным; у них также тормозится воспроизводительная функция. Доминирующие особи стресс-реакции не проявляют. В переуплотненных популяциях стресс распространяется на большую часть особей и, по-видимому, тормозит размножение.
Стрессовое состояние не вызывает необратимых изменений в половой системе, а приводит лишь к временному гормональному блокированию ее функции. После устранения перенаселенности способность к размножению может восстановиться в короткие сроки.
Закономерности стресса, вызываемого перенаселением, изучают в основном на лабораторных животных. Однако многочисленные факты, зарегистрированные в природных популяциях, позволяют предполагать, что в естественных условиях стресс играет немалую роль в регуляции численности и структуры популяций и поведения млекопитающих (рис. 133).
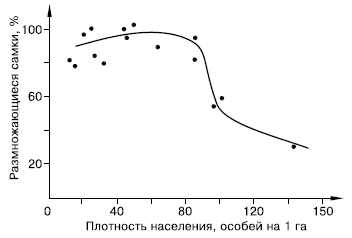
Рис. 133. Зависимость интенсивности размножения от плотности населения в популяции малого суслика (по М. Р. Магомедову, 1995)
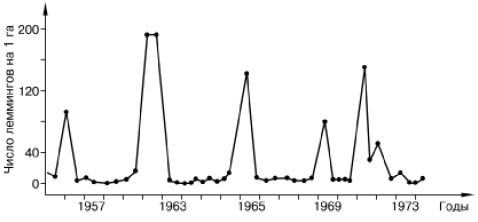
Рис. 134. Динамика численности сибирского лемминга на Аляске (по Bunnel et all., 1975)
Например, в динамике популяций ряда тундровых леммингов зарегистрированы правильные циклы трех-четырехлетней периодичности с амплитудой колебания до 600 крат (рис. 134). Фаза пика в таких колебаниях ограничена обычно одним сезоном, затем следуют резкий спад, фаза депрессии численности и последующего нарастания. При нарастании численности регистрируется увеличение плодовитости, повышение скорости созревания молодняка, усложнение возрастной структуры популяции, снижение смертности новорожденных. В период пика отмечается резкое снижение воспроизводства и одновременно увеличивается смертность во всех возрастных группах. В яичниках самок всех возрастов обнаруживаются серьезные деструктивные изменения, наблюдается массовая гибель фолликулов на ранних стадиях развития. Год-два после спада численности общая интенсивность размножения остается средней, а смертность высокой, а затем вновь все показатели размножаемости начинают расти. У зверьков, родившихся при низкой численности популяции, восстанавливается нормальное функционирование яичников.
На разных этапах этого цикла сильно меняется гормональное состояние зверьков, относящихся к разным генерациям. В пик численности отмечается избыточная активность адреналовой и щитовидной желез, что резко тормозит воспроизводительные функции организма. В ходе цикла у сменяющихся поколений меняется не только функционирование отдельных желез, но и всей эндокринной системы животных. Через несколько поколений, в период минимума численности, состояние эндокринной системы нормализуется и обеспечивает восстановление эффективности процесса размножения. Конкретные причины таких гормональных различий у разных поколений связаны с тем, что жизнеспособность и эндокринные особенности организма формируются в эмбриональный период и во многом определяются физиологическим состоянием родителей.
Таким образом, динамику численности леммингов можно представить как авторегулируемый процесс, в котором большую роль играют эндокринные механизмы.
Все рассмотренные выше примеры взаимодействия между членами популяции, от «жестких» форм – прямого уничтожения одной особью другой – до снижения воспроизводительных способностей как условного рефлекса на повышение частоты контактов, представляют собой разные формы ограничения роста популяций. Эти тормозящие механизмы включаются до полного истощения ресурсов среды в ответ на сигналы, свидетельствующие об угрозе перенаселения.
Степень развития механизмов популяционного гомеостаза находится также в тесной связи с тем, насколько влияют на популяцию другие виды: конкуренты, хищники, паразиты. Общая регуляция численности популяций в природных сообществах происходит в результате сложных межвидовых и внутривидовых взаимоотношений.
Гомеостаз популяцій
— Поддержание определенной численности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит.
Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других — в смягченной, например в понижении плодовитости на основе условных рефлексов.

К жестким формам внутривидовой конкуренции следует отнести, например, явление самоизреживания у растений. При большой густоте всходов часть растений неминуемо погибает в результате угнетения физиологически более сильными соседями. Уменьшение числа растений происходит, даже если высеянные семена генетически однородны. В этом случае, по-видимому, имеют значение разница в размерах, во времени появления всходов, детали микроокружения. В одном из опытов с клевером Trifolium sttb-terraneum через 84 дня после появления всходов на участке в 1 м2 из 1250 растений осталось 650, причем влияние вредителей было исключено.
У райграса многолетнего основной экологической единицей становится не особь, а побег. Обнаружено, что при разных нормах высева семян, от 6 до 180 кг/га, сначала густота побегов варьирует от 30 до 1070 на 100 см2, но затем во всех случаях становится равной примерно 500, т. е. в более редких посевах появляются новые побеги, а в более густых часть отмирает.
Регуляция плотности популяции у растений ввиду особенностей их роста происходит обычно не только путем изменения численности особей на единице площади, но и путем изменения вегетативной мощности каждого. В загущенных посевах растения менее облиственны, с меньшим количеством побегов. Общая их масса при увеличении плотности посевов сначала возрастает пропорционально количеству высеянных семян, а затем остается на постоянном уровне, тогда как средняя масса отдельных особей соответственно уменьшается. В данном случае стабилизируется не число особей в популяции, а общая листовая фотосинтезирующая поверхность растений.
У животных жесткие формы регуляции плотности популяций проявляются обычно лишь в тех случаях, когда запасы пищи, воды или других ресурсов резко ограничены, а животные либо не способны в данный период к поискам ресурсов на другой территории, либо эти поиски неэффективны. Например, в небольших пресноводных водоемах, где нет других видов рыб, популяции окуня могут поддерживать свое существование и регулировать плотность за счет питания взрослых собственной молодью. Мальки же растут за счет мелкого планктона, к питанию которым крупные окуни не приспособлены. Каннибализм возникает иногда в период длительного голодания и в популяциях хищных млекопитающих.
Особый интерес представляют некоторые, сравнительно редкие виды, у которых способность к умерщвлению конкурентов внутри популяции закрепляется эволюционно в их поведении и даже морфологии. Подобные примеры встречаются среди насекомых. Для паразитоидных перепончатокрылых, откладывающих свои яйца в яйца или личинки других насекомых, запас пищи в хозяине весьма ограничен и дает возможность развиться чаще всего лишь одной особи паразита. У ряда видов личинки первого возраста имеют челюсти, ненужные при питании содержимым яйца, но приспособленные для схваток с конкурентами. Нападение личинок друг на друга обычно происходит до того, как они начинают кормиться. После уничтожения конкурента личинка линяет и ее острые челюсти сменяются мягкими и бугорковидными. У тех видов, самки которых способны распознать уже зараженных хозяев и не откладывать яйца в них повторно, личинкам не приходится конкурировать между собой, и они лишены острых челюстей.
Групповые паразиты, откладывающие по нескольку яиц в одно насекомое, нередко обладают способностью соразмерять число яиц с величиной жертвы. Однако при недостатке хозяев возможно перезаражение обнаруженных паразитами, при этом обеспеченность потомства кормом снижается. В таких случаях наблюдается уменьшение размеров личинок. Вышедшие из них имаго откладывают вдвое-втрое меньше яиц при сокращенных сроках жизни. Например, у самых мелких наездников-браконид происходит полная редукция яичников.
Плотность популяции у паразитических перепончатокрылых может регулироваться также через изменения в соотношении полов, что влияет на численность следующего поколения. У многих видов сильно различается смертность самцов и самок при перенаселении. Например, у некоторых браконид доля самок составляет около 70% при слабом заражении насекомых-хозяев и падает ниже 10%—при сильном. Кроме того, соотношение полов регулируется поведением самок. Для многих видов наездников установлено, что в мелких хозяевах, представляющих малый запас корма, самки откладывают преимущественно неоплодотворенные яйца, из которых у перепончатокрылых развиваются самцы. При учащении контактов самок друг с другом или при восприятии следовых запахов, оставляемых другими самками, среди откладываемых яиц также увеличивается доля неоплодотворенных.
Среди механизмов, задерживающих рост популяций, у многих видов большую роль играют химические взаимодействия особей. Так, вода аквариума, в котором содержались дафнии, способна задерживать рост представителей того же вида и сохраняет згу способность в течение нескольких дней. Головастики выделяют в воду частицы белковой природы, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких, так как устойчивость к одной и той же концентрации ингибитора находится в прямой зависимости от размеров. Один крупный головастик Rana pipiens может задержать рост всех других в 75-литровом аквариуме. Поколение, вышедшее в близкие сроки из икры, отложенной в одном водоеме, вскоре разделяется на две размерные группы: более крупные, продолжающие расти головастики и мелкие, затормозившие свой рост из-за неблагоприятной для них концентрации метаболита. Экологическая выгода такого разделения популяции в том, что особи с наследственно более быстрым темпом роста, используя в полной мере кормовые ресурсы водоема, получают возможность быстро завершить метаморфоз и в популяцию вливается полноценное пополнение. Оставшиеся мелкие головастики, после того как первая партия покинет водоем и в нем снизится концентрация ингибитора, также имеют шансы увеличиться в размерах и достичь стадии метаморфоза, но значительно позже. Эту вторую часть пополнения можно рассматривать как своего рода резерв, который вливается в популяцию лишь при достаточно благоприятных условиях (если не пересохнет временный водоем, если в нем продолжится воспроизводство водорослей — основной пищи головастиков и т. п.).

Выделение в окружающую среду продуктов, задерживающих рост, обнаружено у многих растений и водных животных, особенно у рыб.
Другой механизм ограничения численности популяций — такие изменения физиологии и поведения при увеличении плотности, которые в конечном счете приводят к проявлению инстинктов массовой миграции. В результате происходит выселение большей части популяции за пределы территории, занимаемой в оседлый период. Особенно ярко это проявляется у насекомых, которым свойственна фазовость — резко разнокачественное состояние особей в зависимости от плотности популяции. У перелетной саранчи-шистоцерки в постоянных очагах ее обитания в Индии, Пакистане, Восточной Африке и Аравии при низкой численности личинки одиночной фазы имеют ярко-зеленую, а взрослые — серовато-зеленую или бурую окраску. В годы массового размножения, которое наступает при благоприятном сочетании погодных условий, саранча переходит в стадную фазу. Личинки приобретают ярко-желтую, с черными пятнами окраску, взрослая неполовозрелая шистоцер-ка интенсивно розово-лиловая, половозрелая — лимонно-желтая. Изменяется и морфология особей: удлиняются надкрылья, меняются формы переднеспинки, киля, пропорции конечностей и т. п. Переход из одной фазы в другую занимает около трех поколений. Процесс стимулируется зрительным восприятием особей своего вида и контактами при помощи усиков. Это вызывает в организме насекомых целую серию реакций, приводящих к гормональным перестройкам, в которых участвуют эндокринные железы.
Стадная фаза отличается повышенной возбудимостью и чрезвычайной прожорливостью. Плодовитость самок снижается, но они откладывают яйца с большим содержанием питательных веществ. Стадная саранча все время находится в состоянии миграционной активности. Личинки двигаются скоплениями — кулигами, а взрослые гигантскими стаями разлетаются на сотни и тысячи километров от мест постоянного обитания. Так, в конце прошлого столетия масса одной из стай шистоцерки, перелетевшей через Красное море, была определена не менее чем в 44 млн. т.
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2—3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотносга населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поколений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.
Территориальное поведение животных, выработавшееся в ходе эволюции как система инстинктов,—наиболее эффективный механизм сдерживания роста численности популяции на данной площади. Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводят к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства. Это же относится и к выведенному потомству, среди которого лишь некоторая часть вследствие естественной смертности занимает освобождающиеся участки.
Выселения как ответная реакция на растущую плотность популяции свойственны многим видам птиц и млекопитающих. Кроме обычной расселительной дисперсии молодняка, для ряда видов с резкими колебаниями численности характерны массовые перемещения— нашествия. Они возникают нерегулярно, лишь в годы вспышек размножения, и не имеют постоянного направления. Такие нашествия описаны, например, у тундровых леммингов, белок Сибири и Северной Америки и др. При нашествиях часть особей остается на месте, а среди эмигрантов преобладают молодые.
Повышение плотности популяции может сопровождаться такими изменениями в физиологии особей, которые ведут к снижению рождаемости и увеличению смертности. У млекопитающих известно явление стресса, которое впервые было описано в 1936 г. физиологом Г. Селье для человека. В ответ на отрицательное воздействие каких-либо факторов в организме возникают реакции двух типов: 1) специфические, зависящие от природы.повреждающего агента (например, усиление теплопродукции при действии холода), и 2) неспецифическая реакция напряжения как общее усилие организма приспособиться к изменившимся условиям. Эта общая реакция складывается из ряда физиологических и морфологических изменений, которые постепенно развертываются как единый процесс. Реакция напряжения, или стресс, возникает в ответ на любые отрицательные воздействия среды, в том числе и на отклонение плотности популяции от оптимума.
Большую роль в развитии стресса играют сигналы коры головного мозга, меняющие активность гипоталамуса — центрального звена вегетативной нервной системы. В свою очередь, деятельность гипоталамуса вызывает изменения в функционировании ги-пофизарно-надпочечниковой гормональной системы. В состоянии стресса у животных сильно увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов, выделяемых этим органом, а также происходит целый ряд других изменений в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, резорбция эмбрионов, рано прекращается лактация, угасают инстинкты заботы о потомстве и т. п., уменьшается число выводков и количество в них молодых. В конечном счете все это приводит к торможению роста популяции. У мышевидных грызунов, содержащихся в клетках одинаковых размеров, четко проявляется обратно пропорциональная зависимость между численностью зверьков в клетке и массой их репродуктивных органов. В состоянии стресса у животных даже при достаточном снабжении кормом понижается устойчивость к вредным воздействиям среды, что увеличивает смертность.
От плотности населения зависит в первую очередь поведение животных. У многих видов в условиях скученности повышается уровень агрессивности, меняется реакция на особей противоположного пола, молодняк и т. д. Нарушение сложившихся стереотипов поведения приводит большинство особей к состоянию стресса.
Стресс-реакция как механизм, регулирующий рождаемость, особенно отчетливо проявляется у животных с хорошо выраженной системой иерархического подчинения в группах.
Реакция напряжения свойственна подчиненным животным; у них также тормозится воспроизводительная функция. Доминирующие особи стресс-реакции не проявляют. В переуплотненных популяциях стресс распространяется на большую часть особей и, по-видимому, тормозит размножение.
Стрессовое состояние не вызывает необратимых изменений в половой системе, а приводит лишь к временному гормональному блокированию ее функции. После устранения перенаселенности способность к размножению может восстановиться в короткие сроки.

Закономерности стресса, вызываемого перенаселением, изучают в основном на лабораторных животных. Однако многочисленные факты, зарегистрированные в природных популяциях, позволяют предполагать, что в естественных условиях стресс играет немалую роль в регуляции численности и структуры популяций и поведения млекопитающих.
Например, в динамике популяций ряда тундровых леммингов зарегистрированы правильные циклы трех-четырехлетней периодичности с амплитудой колебания до 600 крат. Фаза пика в таких колебаниях ограничена обычно одним сезоном, затем следуют резкий спад, фаза депрессии численности и последующего нарастания. При нарастании численности регистрируется увеличение плодовитости, повышение скорости созревания молодняка, усложнение возрастной структуры популяции, снижение смертности новорожденных. В период пика отмечается резкое снижение воспроизводства и одновременно увеличивается смертность во всех возрастных группах. В яичниках самок всех возрастов обнаруживаются серьезные деструктивные изменения, наблюдается массовая гибель фолликулов на ранних стадиях развития. Год-два после спада численности общая интенсивность размножения остается средней, а смертность высокой, а затем вновь все показателл размножаемости начинают расти. У зверьков, родившихся при низкой численности популяции, восстанавливается нормальное функционирование яичников.
На разных этапах этого цикла сильно меняется гормональное состояние зверьков, относящихся к разным генерациям. В пик численности отмечается избыточная активность адреналовой и щитовидной желез, что резко тормозит воспроизводительные функции организма. В ходе цикла у сменяющихся поколений меняется не только функционирование отдельных желез, но и всей эндокринной системы животных. Через несколько поколений, в период минимума численности, состояние эндокринной системы нормализуется и обеспечивает восстановление эффективности процесса размножения. Конкретные причины таких гормональных различий у разных поколений связаны с тем, что жизнеспособность и эндокринные особенности организма формируются в эмбриональной период и во многом определяются физиологическим состоянием родителей.
Таким образом, регуляцию численности леммингов можно представить как авторегулируемый процесс, в котором большую роль играют эндокринные механизмы.
Все рассмотренные выше примеры взаимодействия между членами популяции, от «жестких» форм — прямого уничтожения одной особью другой — до снижения воспроизводительных способностей как условного рефлекса на повышение частоты контактов, представляют собой разные формы ограничения роста популяций. Эти тормозящие механизмы включаются до полного истощения ресурсов среды в ответ на сигналы, свидетельствующие об угрозе перенаселения.
Степень развития механизмов популяционного гомеостаза находится также в тесной связи с тем, насколько влияют на популяцию другие виды: конкуренты, хищники, паразиты. Общая регуляция численности популяций в природных сообществах происходит в результате сложных межвидовых и внутривидовых взаимоотношений.
Гомеостаз популяции — Студопедия
Популяции организмов обладают способностью к естественному регулированию плотности. Плотность популяции при более или менее значительных колебаниях остается в устойчивом состоянии между своими верхним и нижним пределами. Это обеспечивается действием определенных приспособительных механизмов.
Тенденция популяций поддерживать внутреннюю стабильность с помощью собственных регулирующих механизмов называется гомеостазом, а колебания численности популяций в пределах какой-то средней величины — их динамическим равновесием.Все биологические системы характеризуются способностью к гомеостазу, т.е. к саморегуляции. С помощью саморегуляции поддерживается в целом существование каждой системы — ее состав и структура, внутренние связи и преобразования в пространстве и во времени.
Гомеостатическими являются прежде всего отдельные особи, а затем популяции. Саморегулирующиеся системы не замкнуты, они активно взаимодействуют с внешней средой и, следовательно, подвержены изменениям. Саморегуляция — необходимое приспособление организмов для поддержания жизни в постоянно меняющихся условиях. Саморегулирование популяции осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами. С одной стороны, это биотический потенциал, составляющий совокупность всех факторов, способствующих увеличению численности популяции, а с другой — это сопротивление среды — совокупность факторов, снижающих численность популяции (рис. 16).
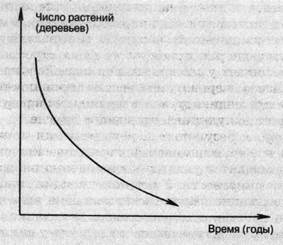
Рис. 17. Ход самоизреживания леса с возрастом
К механизмам внутрипопуляционного гомеостаза относятся угнетающие (ингибирующие) выделения во внешнюю среду более сильными особями, стрессовые явления, территориальность, миграции между популяциями. Выделения во внешнюю среду характерны как для растительных, так и для животных организмов. На лабораторных животных (крысы, мыши) показано, что перекачивание воздуха из перенаселенных помещений в те помещения, где животные свободно размножаются и нормально развиваются, приводит к замедлению роста и угнетению этих животных. Аналогичные результаты наблюдались в опытах с головастиками лягушек, когда в аквариум, где они хорошо развивались, добавляли воду из аквариумов, в которых плотность организмов была чрезвычайно высокой.
Явления территориальности наиболее четко выражены в животном мире. К ним относятся различные способы охраны занимаемых территорий. У кошачьих и tсобачьих территориальность проявляется через мечение границ участков выделением желез, мочой или механическими отметинами на деревьях и почве. Пение птиц — это прежде всего сигнал о занятости территории в период размножения и последующего выкармливания потомства.
Важным механизмом регуляции численности, проявляющимся в переуплотненной популяции, является стресс-ре-
акция (от англ. stress — напряжение). Если на популяцию воздействует какой-то сильный раздражитель, она отвечает на него неспецифической реакцией, называемой стрессом. В живой природе различают много форм стресса: антропи-ческий (возникает у животных под воздействием деятельности человека), нервно-психический (проявляется при несовместимости индивидуумов в группе или в результате переуплотнения популяции), шумовой и другие.
Например, в результате переуплотнения популяции, у отдельных особей возникают физиологические изменения, которые приводят к резкому сокращению рождаемости и увеличению смертности. У млекопитающих такое явление получило название стресс-синдрома. При этом животные становятся настолько агрессивными (жестокие драки, нетерпимость присутствия соседа и т.д.), что у них почти полностью прекращается размножение. У животных, находящихся в стрессовом состоянии, увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов. У самок нарушается овуляция, происходит резорбция эмбрионов, не проявляются инстинкты заботы о потомстве и т.д.
Миграции, как фактор гомеостаза могут проявляться в двух видах. Первый относится к массовому исходу особей из популяции при явлениях перенаселенности (нашествие). Такие явления характерны для леммингов, белок и других видов с взрывным типом динамики численности. Второй вид миграции связан с более постепенным (спокойным) уходом части особей в другие популяции с меньшей плотностью населения.
Гомеостаз в полной мере проявляется, если срабатывают все механизмы, лежащие в его основе. В настоящее время нарушения гомеостатических механизмов вызываются в большинстве случаев антропогенными факторами. В связи с этим, одной из важнейших задач человека является исключение или резкое снижение действия подобных факторов.
Общие принципы популяционного гомеостаза
То же можно сказать и о таких особенностях популяций, как средние размеры индивидуальных (или стадных) территорий, степень подвижности животных и т. п. Упоминавшаяся в гл. 10 сезонная динамика пространственной структуры у рада видов — пример устойчивого приспособления к закономерно повторяющейся смене условий жизни популяции. Характерным свойством популяции может быть и средний уровень плодовитости. Адаптивный характер этого показателя может проявляться в увеличении плодовитости в популяциях, занимающих пессимальные местообитания, что компенсирует повышенную смертность в этих условиях.[ …]
Механизмы формирования и закрепления уровня стабилизации наиболее генеральных популяционных свойств основываются в первую очередь на генетических (связанных с определенной степенью изоляции) и микроэволюционных процессах.[ …]
Изменчивость, динамичность конкретных условий жизни вызывают формирование лабильных функциональных адаптаций, действующих на данном уровне стабилизации популяционных функций и поддерживающих этот уровень. Механизмы таких адаптаций основаны на эколого-физиологических процессах. При всем многообразии конкретных проявлений лабильные адаптации популяционного уровня действуют по единому принципу обратной связи: в ответ на внешние или внутрипопуляционные изменения они вызывают адекватные сдвиги во внугрипопуляционных отношениях, продолжающиеся до тех пор, пока не восстановятся «уравновешенные» отношения между популяцией и средой. Именно лабильные авторегуляторные процессы придают определенную устойчивость популяционным системам на фоне изменчивых условий их жизни.[ …]
В том случае, когда изменения среды выходят за пределы адаптированного диапазона, биологически выгодной оказывается смена уровня стабилизации системы. В этом случае на основе внутри-популяционной разнокачественности начинаются процессы отбора, ведущие к перестройке наиболее генерализованных качеств популяции в соответствии с изменившимися средними условиями ее жизни. Таким образом, гомеостатические механизмы популяционного уровня лежат в основе того, что в относительно стабильных условиях популяция функционирует как устойчивая форма существования вида, а в условиях направленного изменения среды представляет собой исходную элементарную единицу эволюционного процесса.[ …]
Гомеостазирование популяционных функций связано с такими показателями, как численность и плотность населения. Это положение исходно предусматривает динамический характер основных популяционных параметров: Как численность популяции, так и ее выражение, отнесенное к единице пространства, не остается одинаковым во времени. Свойственные популяции процессы постоянного воспроизведения сопровождаются столь же постоянным отмиранием особей. В силу многих причин не остается неизменной и возможность заселения отдельных элементов ландшафта: «емкость угодий» меняется в сезонном и многолетнем масштабе, что определяет динамику параметров плотности населения даже при постоянном уровне репродукции. В конкретных популяциях, локализованных в определенных границах пространства, постоянно происходят процессы притока особей извне и выселения определенной части населения за пределы популяции. Все эти процессы детерминируют пульсирующий, динамический характер популяции как системы, составленной множеством отдельных организмов. Будучи представителями одного вида, организмы в составе популяции несут рад общих признаков, определяющих биологическую специфику популяции в целом и ее место в функционировании глобальных систем биологического круговорота. В то же время составляющие популяцию особи отличаются друг от друга по полу, возрасту, генетическим особенностям и месту в функциональной структуре популяции. На этой основе формируются такие фундаментальные популяционные процессы, как типы воспроизводства и смертности, масштабы и формы пространственной дисперсии, общий уровень биологической активности и др.[ …]
Вернуться к оглавлению
Гомеостаз зависит от механизмов регуляции!
Ресурс исследования
- Проводить исследования
- Искусство и гуманитарные науки
- Бизнес
- Инженерная технология
- Иностранный язык
- история
- математический
- Наука
- Социальная наука
Лучшие подкатегории
- Продвинутая математика
- Алгебра
- Базовая математика
- Исчисление
- Геометрия
- Линейная алгебра
- Предалгебра
- Предварительный расчет
- Статистика и вероятность
- Тригонометрия
- другое →
Лучшие подкатегории
- Астрономия
- Астрофизика
- Биология
- Химия
- Науки о Земле
- Наука об окружающей среде
- Науки о здоровье
- Физика
- другое →
Лучшие подкатегории
- Антропология
- Закон
- Политология
- Психология
- Социология
- другое →
Лучшие подкатегории
- Бухгалтерский учет
- Экономика
- Финансы
- Менеджмент
- другое →
Лучшие подкатегории
- Аэрокосмическая техника
- Биоинженерия
- Химическая инженерия
- Гражданское строительство
- Компьютерные науки
- Электротехника
- Промышленное проектирование
- Машиностроение
- Веб-дизайн
- другое →
Лучшие подкатегории
- Архитектура
- Связь
- Английский
- Гендерные исследования
- Музыка
- Исполнительское искусство
- Философия
- Религиоведение
- Письмо
- другое →
Лучшие подкатегории
- Древняя история
- Европейская история
- История США
- Всемирная история
- другое →
.
Секс и половые гормоны в гомеостазе тканей
1. Введение
Живые системы постоянно подвергаются воздействию потенциально токсичных внутренних и внешних процессов. Нормальная метаболическая функция клеток производит множество потенциально повреждающих окислительных метаболитов, вызывающих повреждение ДНК, белков и липидов. Кроме того, живые клетки подвергаются воздействию множества внешних факторов, которые могут быть интернализованы как строительные блоки и / или источники энергии. Эти жизненно важные процессы ставят организм под угрозу.Чтобы избежать повреждений, необходимы стратегии преодоления трудностей. Существует несколько линий клеточной защиты, индуцируемой путями клеточного стресса, включая процессы компартментализации, ферментативную модификацию, экстернализацию, деградацию и репарацию [1, 2].
В конечном счете, этих процессов может быть недостаточно для предотвращения серьезного повреждения клеток. Следовательно, каждая клетка дополнительно оснащена программами внутренней гибели клеток, которые можно активировать, чтобы предотвратить нанесение поврежденной клеткой вреда организму [3].Потери клеток неизбежны и происходят в нашем организме непрерывно, даже без специального триггера. Скорость гибели клеток может быть значительно увеличена во время более серьезных проблем. Клеточные потери необходимы для предотвращения пагубных эффектов, таких как трансформация опухоли [4]. Таким образом, запрограммированная гибель клеток должна быть тщательно сбалансирована, и эта способность является ключевым фактором, определяющим здоровье и выживание организма.
Клетки, потерянные в результате гибели клеток, необходимо заменить для поддержания количества клеток и, в конечном итоге, функции тканей.Таким образом, контролируемая регенерация необходима для того, чтобы справиться с потерями клеток из-за токсических воздействий, происходящих из внутренних и внешних источников, независимо от того, являются ли они результатом нормальных метаболических процессов или повреждающих стимулов окружающей среды. Потери и пролиферацию клеток необходимо тщательно сбалансировать, чтобы гарантировать надлежащее функционирование. Таким образом, процесс тканевого гомеостаза, то есть способность направлять поврежденные клетки в программы гибели клеток, заменять клетки путем пролиферации и регулировать точный баланс этих событий, является критически важным процессом для сохранения здорового состояния.Любое нарушение баланса между гибелью клеток и их пролиферацией — будь то подавляющие повреждающие события за пределами диапазона толерантности хозяина и / или в первую очередь неэффективные или дезадаптивные гомеостатические механизмы хозяина — склонно вызывать сбои в работе органов в организме, в конечном итоге вызывая болезнь и потенциально смерть.
2. Половые и половые гормоны в гомеостазе тканей
2.1. Происхождение половых различий
В процессе развития половые различия возникают в результате генетических и гормональных влияний.Главные регуляторы дифференциации мужского пола, такие как SRY, кодируются ДНК Y-хромосомы, управляющей эмбриональным развитием мужского фенотипа у млекопитающих. Развитие гонад у женщин или мужчин вызывает гормональную среду, специфичную для пола [5]. Половые гормоны в течение всей жизни вызывают организационные эффекты, вызывая стойкие половые изменения в тканях, например, посредством эпигенетических модификаций [6, 7]. Активационные эффекты дополнительно вносят половые различия в структуру и функции тканей в зависимости от характера воздействия гонадных гормонов.На протяжении всей жизни поражаются все органы тела [8]. В этом отношении половые различия основаны на различном хромосомном оборудовании, которое определяет каждую клетку тела как мужскую или женскую. Эти основные различия в дальнейшем формируются половыми гормонами в зависимости от предыдущего или текущего, временного или постоянного воздействия [9]. Эта гипотеза была сформулирована Арнольдом [10] следующим образом: «XX и XY клетки различаются до секреции гонадных гормонов, и гонадные гормоны влияют на XX и XY клетки неодинаково.”
Половые гормоны действуют через рецепторы, широко экспрессирующиеся во всех клетках тела. Классические рецепторы эстрогена (ERα, ERβ), андрогена (AR) и прогестерона (PR) принадлежат к семейству белков ядерного рецептора, действующего через ядро как фактор транскрипции или кофактор. В дополнение к их ядерному действию было обнаружено, что они локализованы на клеточной мембране и митохондриях, индуцируя быстрые негеномные внутриклеточные сигнальные пути, например, посредством взаимодействия с фактором роста или рецепторами цитокинов.Эстроген-связывающие клеточные рецепторы, локализованные на мембране из семи семейств трансмембранных рецепторов, также были охарактеризованы, например, рецептор эстрогена, связанный с G-белком, GPR30 / GPER-1 [11, 12, 13].
В конечном счете, различия между полами происходят из хромосомных и гормональных половых различий, на которые в дальнейшем влияют факторы окружающей среды. Таким образом, различия, происходящие от биологического пола, в дальнейшем определяются полом, который относится к восприятию мужской или женской идентичности и зависит от социальных структур на основе пола [8].
2.2. Влияние половых и половых гормонов на пролиферацию клеток
Тканевый гомеостаз гарантирован, когда клетки, потерянные в физиологическом тканевом обороте или в стрессовых условиях, заменяются пролиферацией. Органы с высокими требованиями к функциям обладают повышенным потенциалом регенерации и постоянно обновляют свои клеточные популяции. Это, например, касается кишечника, кожи и крови. Печень, кость и кровь даже могут полностью восстановиться до исходного размера после потери ткани [14].Другие органы имеют более низкий регенеративный потенциал, например сердце, мозг и почки [15, 16, 17]. Многие органы содержат ниши стволовых клеток, в которых находятся стволовые клетки взрослой ткани, которые являются клетками-предшественниками, поддерживаемыми в относительно недифференцированном состоянии, готовыми заменить потерянные клетки путем пролиферации с последующей дифференцировкой [18, 19].
Половые гормоны классически участвовали в регуляции пролиферации клеток репродуктивных органов и рака репродуктивной ткани [20, 21, 22, 23, 24].Помимо этих эффектов, половые гормоны также оказывают заметное влияние на пролиферацию различных популяций стволовых клеток. Было обнаружено, что пролиферация эмбриональных стволовых (ES) клеток усиливается за счет женских гонадных гормонов [25]. ES-клетки происходят из внутренней клеточной массы эмбриоидного тела. Они могут самообновляться in vitro и являются плюрипотентными, то есть могут дифференцироваться во все типы клеток организма [26]. Эстроген, по-видимому, действует через сигнальные пути ядра и клеточной поверхности, включая активацию Erk1 / 2, циклин-зависимые киназы и протоонкогены, такие как c-myc, c-fos, c-jun и pRB в ES-клетках.Кроме того, было обнаружено, что кальциевые каналы, управляемые магазином, играют роль в эстроген-опосредованной пролиферации клеток посредством фактора транскрипции NF-AT [27].
Было также показано, что эстроген влияет на дифференцировку ES-клеток в дофаминергические нейроны. ERβ способствовал дифференцировке путем перекрестной передачи сигналов с инсулиноподобным фактором роста-1 [28]. Было обнаружено, что дифференцировка моторных нейронов от ES-клеток усиливается 17-β эстрадиолом и прогестероном через ядерный ERα и рецептор прогестерона [29].Было обнаружено, что дофаминергические предшественники, полученные из ES-клеток, увеличивают пролиферацию при лечении прогестероном in vitro [30].
Индуцированные плюрипотентные стволовые (iPS) клетки похожи на ES-клетки в отношении их способности дифференцироваться во все типы клеток, что является многообещающим инструментом для исследований in vitro и регенеративной медицины. Они происходят из зрелых клеток взрослых путем перепрограммирования путем введения специфических факторов транскрипции [31]. Было показано, что половые гормоны, как и ES-клетки, влияют на iPS-клетки.Нейрональные клетки, полученные в результате дифференцировки iPS-клеток, показали усиление ветвления дендритов при обработке 17-β эстрадиолом [32]. Было обнаружено, что функциональная интеграция дофаминергических нейрональных клеток из iPS-клеток в нейрональные цепи усиливается эстрадиолом [33]. Было описано, что тестостерон усиливает дифференцировку iPS-клеток в инсулин-продуцирующие клетки [34].
Половые различия также описаны для тканевых стволовых клеток in vivo. Считается, что взрослые стволовые клетки обеспечивают локальный пул самообновляющихся мультипотентных клеток, играющих ключевую роль в гомеостазе тканей и восстановлении после повреждения [35].Стволовые клетки во многих нишах стволовых клеток, по-видимому, обладают более высокой способностью к самообновлению, обладают повышенным потенциалом регенерации и в некоторых случаях проявляют более высокую пролиферативную активность у женщин [36, 37]. Внутренний половой диморфизм был описан для нервных стволовых клеток, которые открывают большие перспективы для лечения потенциальных повреждений мозга в будущем. Например, было показано, что пролиферация нервных стволовых клеток зависит от гормональных изменений у взрослых мышей из-за эстрального цикла, беременности, репродуктивного статуса и возраста.Было обнаружено, что фазы высокого воздействия эстрогенов, такие как проэструс, связаны с усилением нейрогенеза в гиппокампе у взрослых, что указывает на роль эстрогенов [38]. Дифференциальная экспрессия рецепторов половых стероидов и ферментов, метаболизирующих андрогены, может привести к разным результатам при трансплантации нервных стволовых клеток [39]. Было обнаружено, что пролиферация нервных стволовых клеток зависит от ядерных ER, тогда как дифференцировка олигодендроглии стимулировалась ERs, ассоциированными с клеточной мембраной [40]. Другой исследователь также предположил, что действие половых стероидов на мозг может быть связано с уменьшением повреждения мозга.Было обнаружено, что интактные самки менее подвержены травмам, чем самки и самцы после овариэктомии [41]. Точно так же стволовые клетки мышечного происхождения, полученные от самок мышей и трансплантированные дистрофическим мутантным мышам, показали лучший потенциал для регенерации скелетных мышц, чем стволовые клетки самцов [42].
Было обнаружено, что гемопоэтические стволовые клетки более многочисленны и пролиферативны у самок мышей по сравнению с самцами, зависимыми от воздействия эстрогена [43]. Было обнаружено, что 17-β эстрадиол улучшает гематопоэтическую дифференцировку из человеческих iPS-клеток и из человеческой пуповинной крови посредством передачи сигналов ERα, что указывает на универсальную функцию эстрогена в дифференцировке гемопоэтических стволовых клеток [44, 45].
Эстрогены благотворно влияют на регенерацию костей [46]. Эстроген стимулирует пролиферацию остеобластов с различными ролями для ERα и ERβ [47]. In vitro было обнаружено, что пролиферация мезенхимальных стромальных клеток костного мозга усиливается эстрогеном [48]. Эстрогены усиливают пролиферацию и миграцию эндотелиальных клеток-предшественников костного мозга в ишемические области сердца, способствуя восстановлению и регенерации [49]. Было также описано, что андрогены стимулируют пролиферацию и способность к ангиогенезу / восстановлению сосудов циркулирующих эндотелиальных клеток-предшественников у мужчин, а не у женщин [50].
2.3. Половые различия в выживаемости клеток
Когда клетки-предшественники, участвующие в регенерации ткани, входят в состояние клеточного старения, гомеостаз ткани может быть нарушен. Клетки способны навсегда останавливать клеточный цикл и оставаться в спокойном, но все же функциональном состоянии [51]. Это возможная судьба клеток, поврежденных без возможности восстановления. Три основных типа старения — это репликативное старение, старение, вызванное онкогенами, и повреждение ДНК, вызванное повреждением ДНК. Путь ответа на повреждение ДНК, по-видимому, в конечном итоге участвует в выполнении программы независимо от первичного стимула [52].Кроме того, стареющие клетки способны влиять на свои соседние клетки путем секреции ряда активирующих сигналов, называемых секреторным фенотипом, связанным со старением. Сигналы могут способствовать провоспалительному или — альтернативно — иммуносупрессивному / профибротическому состоянию. Обе фазы, по-видимому, важны для успешного восстановления тканей, и время сдвига секретома может быть решающим [53]. Было показано, что этиология и прогрессирование многих раковых или возрастных заболеваний зависит от секретома стареющих клеток [54, 55, 56].
В качестве альтернативы, клетки могут активировать программу клеточной гибели как средство обеспечения физиологического обновления ткани или в ответ на сильное повреждение. Наиболее распространены программы гибели клеток I типа или апоптоз, тип II или аутофагия, тип III или некроз и митотическая катастрофа [57, 58]. Методы гибели клеток характеризуются различными морфологическими критериями и выполняются с помощью определенных внутриклеточных сигнальных каскадов. Специфические катаболические ферменты обычно связаны с определенными формами гибели клеток, например, каспазы с апоптозом.Пути взаимозависимы. Интенсивность сигнала повреждения часто имеет решающее значение для типа выполняемой программы гибели клеток или переключения с одной модальности на другую. Кроме того, аутофагия в первую очередь не рассматривается как механизм гибели клеток. Аутофагия описывает процесс, связанный с разрушением и переработкой определенных субклеточных органелл. Этот процесс может обеспечить стратегию выживания клеток за счет уменьшения поврежденных органелл и / или сдвига внутренних ресурсов с целью оптимизации выживания клеток.Только если повреждающий процесс превышает клеточную защиту, клетки погибают в процессе [59, 60].
Что касается роли пола в принятии решений о судьбе клеток, то в нескольких отчетах были выявлены отчетливые полозависимые различия. Было показано, что половые гормоны влияют на склонность клеток к апоптозу. В целом было обнаружено, что более низкие концентрации эстрогена обладают защитным действием, в то время как более высокие концентрации способствуют апоптозу. Было обнаружено, что андрогены усиливают, но также подавляют апоптоз в зависимости от клеточного контекста [61, 62].Например, было описано, что эстроген и тестостерон уменьшают апоптоз в клетках скелетных мышц [63]. Оба гормона, по-видимому, также предотвращают апоптоз нейрональных клеток, усиливая их нейропротекторную функцию [64, 65]. Антиапоптотическое действие тестостерона также было описано в β-клетках поджелудочной железы самцов крыс, но не самок крыс [66]. Однако эстроген и эстрогенные соединения, по-видимому, усиливают апоптоз β-клеток поджелудочной железы у пожилых мышей и снижают апоптоз у молодых животных [67].Что касается эндотелиальных клеток сосудов, несколько исследований показали, что эстрогены защищают от апоптотической гибели клеток [68, 69], тогда как апоптоз увеличивается в эндотелии коронарных артерий у женщин в постменопаузе [70]. Было обнаружено, что тестостерон вызывает апоптоз эндотелиальных клеток [71, 72, 73]. Лечение тестостероном также индуцировало апоптоз или старение в клетках кожных сосочков человека — процесс, характерный для наследственной мужской алопеции [74, 75]. Кроме того, было обнаружено, что андрогены способствуют апоптозу клеточных линий почек и кишечника, а также макрофагов, происходящих из костного мозга [76, 77, 78].
В целом, половые гормоны, по-видимому, влияют на решения клеточной судьбы в зависимости от клеточного контекста. Также очевидны гормонально-независимые половые различия, формирующие клеточный ответ [79]. Таким образом, женские и мужские клетки, по-видимому, полагаются на разные стратегии преодоления стресса. Например, гладкомышечные клетки сосудов, выделенные из аорты самцов крыс, более склонны к апоптозу в ответ на УФ-облучение, тогда как женские клетки более склонны выполнять программу клеточного старения [62, 80].Женские клетки продемонстрировали характеристики аутофагии, которая, как предполагается, помогает женским клеткам восстанавливать вызванные ультрафиолетом внутриклеточные повреждения, в конечном итоге обеспечивая стратегию выживания [81]. Кроме того, было обнаружено, что женские клетки лучше прилипают к опоре роста, что позволяет избежать инициации апоптотической гибели клеток за счет отслоения клеток, процесса, называемого аноикис-устойчивостью. Различия во внутриклеточной организации актинового цитоскелета и повышенное фосфорилирование киназы фокальной адгезии были приписаны этой более высокой склонности женских клеток к прикреплению [81].По-видимому, женские клетки лучше оснащены для предотвращения гибели клеток. Хотя было обнаружено, что аутофагические процессы защищают нейрональные клетки от гибели клеток из-за голода у самок крыс, мужские клетки не могли получить пользу и чаще умирали от гибели аутофагических клеток [82]. Специфические для органов половые различия были обнаружены в конститутивной аутофагии, процессе, участвующем в физиологическом обновлении тканей. Хотя аутофагические маркерные белки были увеличены в сердце и печени у мужчин по сравнению с женщинами, в почках таких различий не наблюдалось [83].Остеобласты показали снижение аутофагии у стареющих самок мышей, в то время как у самцов показатель оставался постоянным на протяжении всей жизни. Это коррелировало с более высоким окислительным стрессом в женских клетках, таким образом потенциально увеличивая потерю костной массы и играя роль в патофизиологии остеопороза у женщин [84]. Эстрогены изменяют окислительно-восстановительный баланс и противодействуют потере костной массы [46]. Было обнаружено, что на стволовые клетки, участвующие в образовании остеобластов, а именно, мезенхимальные стволовые клетки костного мозга, влияют эстрогены, которые не только вызывают повышенную пролиферацию, но также снижают старение и апоптоз [85].
В целом, повышенная антиоксидантная клеточная защита была вовлечена в обеспечение самок лучшими стратегиями борьбы с окислительным стрессом и предотвращения потери клеток [62]. Различия в базальном окислительно-восстановительном состоянии и реакции на окислительный дисбаланс были продемонстрированы между женскими и мужскими клетками [86]. Например, было показано, что женские клетки производят меньше перекиси водорода и супероксид-аниона. Антиоксидантные ферменты, такие как супероксиддисмутаза (СОД) и каталаза, показали более высокую основную активность в женских клетках по сравнению с мужскими [87].Тиоредоксинредуктазы и СОД марганца повышались эстрогеном в кардиомиоцитах [88, 89]. В гладкомышечных клетках сосудов и циркулирующих моноцитах эстроген стимулирует экспрессию марганца и внеклеточной SOD [90]. Кроме того, было показано, что эстроген по-разному модулирует экспрессию других ключевых ферментов молекулярной защиты в клетках XX и XY, например, поли-АДФ-рибозо-полимераза (PARP), фермент восстановления повреждений ДНК или RLIP76, белок-переносчик, защищающий клетки. [86].
PARP также играет важную роль в половых различиях при инсульте.Эксперименты на мышах показали, что ишемическая гибель нейрональных клеток зависит от передачи сигналов интактной нейрональной синтазы оксида азота (nNOS) / PARP, в то время как у самок защита обеспечивается эстрогеном, что парадоксальным образом также требует интактной оси nNOS / PAPR [91]. В то время как мужские нейрональные клетки, по-видимому, умирают посредством PARP-опосредованного каспазно-независимого пути, пути ишемической гибели клеток, по-видимому, зависят от активации каспаззависимых путей гибели клеток у женщин [9, 92]. Такие половые различия могут иметь отношение к половым различиям в распространенности инсульта [93, 94].
Передача сигналов
PARP также подразумевается в половых различиях в решениях клеточной судьбы в клетках почек. В модели иммуноопосредованного нефрита на мышах передача сигналов PARP индуцировала некроз в мужских клетках и ингибирование PARP сдвигало гибель провоспалительных некротических клеток на противовоспалительный путь апоптоза. В женских клетках, напротив, клеточная гибель не зависела от PARP, и женские клетки предпочтительно подвергались апоптозу. Эстроген действует на выживание только в женских клетках. Помимо клеток почек, гемопоэтические клетки костного мозга показали сходные половые различия [95].
Митохондрии играют решающую роль в программах апоптотической гибели клеток. Было описано, что эстрогены модулируют склонность митохондрий к инициации апоптоза [61, 96]. Эстроген-опосредованная модуляция функции митохондрий достигается воздействием гормонов на экспрессию митохондриальных белков, кодируемых ядерным геномом [97, 98, 99, 100, 101]. Поскольку митохондрии играют центральную роль в защите клетки от окислительного стресса, митохондрии особенно чувствительны к накоплению повреждений с течением времени.Сбои в работе митохондрий накапливаются во время старения, и этот процесс считается одним из основных факторов возникновения многих возрастных заболеваний [102, 103]. В этом процессе наблюдались половые различия. Отсроченное нарушение работы митохондрий во время процесса старения может предоставить женщинам лучшие стратегии борьбы с клеточными стрессорами. Передача митохондрий от матери, по-видимому, обеспечивает более благоприятную среду для потомства женского пола [104]. Xist, ген, кодирующий РНК, участвующий в инактивации Х-хромосомы в женских клетках, по-видимому, имеет решающее значение для поддержания митохондрий [105].Биогенез и деградация митохондрий в результате митофагии зависят от факторов транскрипции p53 и FOXO [106, 107]. Сообщалось о половых различиях в активности этих ядерных факторов. Было показано, что самцы проявляют относительно большую активность FOXO. У женщин, с другой стороны, была более высокая активность p53, что приводило к половозависимым различиям в способности поддерживать здоровую митохондриальную функциональность во время старения [105].
2.4. Возможные последствия специфичных для пола различий в гомеостазе тканей
Вышеупомянутые параграфы описали примеры специфичных для пола различий в отношении процессов, участвующих в поддержании тканей, таких как контроль пролиферации и гибели клеток.Такие эффекты могут в конечном итоге привести к различиям в способности женских и мужских тканей справляться со стрессорами, влияющими на способность восстанавливать и восстанавливать функции или развивать заболевание. Многие заболевания демонстрируют разные уровни заболеваемости и распространенности у мужчин и женщин, что обусловлено патофизиологическими механизмами, специфичными для пола и пола. Половые и гендерные различия интенсивно изучались, в частности, в нервной системе, сердечно-сосудистой системе и развитии рака [108, 109, 110].
Механизмы, лежащие в основе различий в заболеваниях почек между мужчинами и женщинами, менее известны, несмотря на то, что почечные заболевания с высоким риском заболеваемости и смертности являются сложной проблемой для пациентов, клиницистов и общества [111, 112].Международные регистры показывают, что почечная недостаточность развивается у женщин реже, чем у мужчин [113, 114, 115, 116]. Однако основные причины широко неизвестны. Предполагаемые женские защитные эффекты наиболее выражены у женщин репродуктивного возраста [117, 118, 119, 120]. Это открытие предполагает, что женские половые гормоны могут играть ключевую роль. Было высказано предположение, что эстроген обладает ренопротекторным действием за счет регулирования перфузии почек и воздействия на сосудистую сеть. Кроме того, была предложена роль эстрогена в контроле местной ренин-ангиотензиновой системы почек [121, 122, 123].С другой стороны, эстроген участвует в контроле пролиферации мезангиальных и канальцевых клеток и связан с неопластической трансформацией почек в почках хомяка. Было показано, что низкие концентрации эстрогенов вызывают пролиферацию мезангиальных клеток клубочков, в то время как высокие концентрации подавляют ее [124]. Было показано, что первичные эксплантаты клеток проксимальных канальцев и субкультивированные диссоциированные клетки проксимальных канальцев пролиферируют при обработке эстрогеном в физиологических концентрациях [125]. Это открытие было подтверждено на первичных клетках проксимальных канальцев кролика, которые показали повышенную пролиферацию после лечения эстрогенами [126].
Ранее мы показали, что белки, специфичные для клеток почечных канальцев, чаще появляются в моче здоровых женщин на определенных фазах гормонального перехода естественного овулирующего менструального цикла. Образцы мочи здоровых пробандов показали повышенную скорость экскреции с мочой маркерных белков фруктозо-1,6-бисфосфатазы и глутатион-S-трансферазы α, когда уровни эстрогена снизились после предыдущего роста, связанного с овуляцией и лютеиновой фазой [127]. Оба фермента специфически обнаруживаются в клетках проксимальных канальцев, наиболее густонаселенном типе клеток почек.Когда клетки проксимальных канальцев повреждены, внутриклеточные ферменты выделяются в мочу, что делает их клиническими маркерами повреждения почек. В отличие от женщин с овуляцией, пробанды-мужчины и женщины в постменопаузе с течением времени демонстрировали стабильно низкие уровни этих белков-маркеров почек. Другие белки мочи, например, альбумин, α1-микроглобулин и иммуноглобулин G, которые являются маркерами функциональных изменений клубочкового фильтра и / или резорбции канальцевых белков, показали постоянную экскрецию с мочой, что позволяет предположить, что наблюдаемое увеличение высвобождения белка-маркера проксимальных канальцев в овуляция у женщин не сопровождается серьезным нарушением функции почек [127].Этот паттерн экскреции белков-маркеров с мочой предполагает, что циклические изменения женских гормонов могут влиять на здоровье клеток почек. Тубулярные ферменты выделяются в мочу, если клетки проксимальных канальцев отслаиваются и / или их плазматические мембраны становятся негерметичными. Это может быть связано с тем, что канальцевые клетки временно более склонны к повреждению in situ, что приводит к утечке плазматической мембраны, или к тому, что клетки удаляются из канальцевого эпителия и высвобождаются в мочевое пространство, например, в результате апоптоза.Оба процесса приводят к увеличению потерь клеток. Тканевый гомеостаз будет поддерживаться, если повышенное удаление клеток сопровождается повышенной пролиферацией клеток. Это может иметь место во время фаз высокого воздействия эстрогена, предшествующих наблюдаемому выделению канальцевых ферментов в мочу. Открытие того, что тубулярные клетки способны пролиферировать при лечении эстрогенами [126], согласуется с этой гипотезой. Такое периодическое взаимодействие между пролиферацией и потерей клеток, вызванное специфическими изменениями в характере воздействия половых гормонов, может привести к увеличению скорости обновления тканей.Если бы это было так, то женщины репродуктивного возраста обладали бы эффективными средствами, позволяющими легко избавиться от потенциально травмированных, дисфункциональных или просто старых проксимальных канальцевых клеток, заменяя их свежими новыми клетками. Такая временно повышенная репарационная способность может обеспечить эффективное средство для циклического обновления ткани почечных канальцев, что приводит к более высокой устойчивости к повреждению. Однако также возможно, что во время коротких фаз повышенной гибели канальцевых клеток почки могут быть особенно чувствительны к повреждению.Что касается потенциального полезного действия лечения клеток проксимальных канальцев почек фактором роста, индуцирующим пролиферацию, мы ранее продемонстрировали, что лечение эпидермальным фактором роста (EGF) способно ускорять восстановление тканей после лечения интерфероном α (IFNα) in vitro. , Однако, если EGF присутствовал до или во время лечения IFNα, дестабилизация эпителиального барьера усиливалась [128, 129]. Следовательно, общий эффект может быть другим в контексте других циклов или при гормональной терапии, если уязвимые фазы не могут быть ограничены короткими периодами.
.
Влияние и основные механизмы биологически активных соединений на сахарный диабет 2 типа и болезнь Альцгеймера
Сахарный диабет 2 типа — сложное нарушение обмена веществ, характеризующееся гипергликемией и непереносимостью глюкозы. Болезнь Альцгеймера — это прогрессирующее заболевание головного мозга, характеризующееся хронической потерей когнитивных и поведенческих функций. Учитывая общие характеристики обоих заболеваний, общие терапевтические и профилактические средства могут быть эффективными. Биоактивные соединения, такие как полифенолы, витамины и каротиноиды, содержащиеся в овощах и фруктах, могут оказывать антиоксидантное и противовоспалительное действие.Эти эффекты делают их подходящими кандидатами для профилактики или лечения диабета и болезни Альцгеймера. Все больше данных, полученных на клеточных или животных моделях, предполагают, что биоактивные соединения могут оказывать прямое воздействие на уменьшение гипергликемии, усиление секреции инсулина и предотвращение образования амилоидных бляшек. Возможные лежащие в основе молекулярные механизмы описаны в этом обзоре. Необходимы дополнительные исследования для установления клинических эффектов биологически активных соединений.
1. Введение
Диабет — это сложное метаболическое нарушение, которое характеризуется гипергликемией из-за инсулиновой недостаточности и / или дисфункции инсулина.В 2017 году в мире насчитывалось 425 миллионов взрослых людей, страдающих сахарным диабетом. Согласно прогнозам, к 2045 году это число возрастет до 629 миллионов диабетиков во всем мире [1]. При диабете 1 типа гипергликемия вызывается аутоиммунным разрушением бета-клеток поджелудочной железы [2]. Сахарный диабет 2 типа (СД2) является более распространенным типом диабета, при котором периферическая инсулинорезистентность и компенсаторная повышенная секреция инсулина могут ускорять снижение секреторной функции островков поджелудочной железы, что в конечном итоге приводит к дефициту инсулина [2].Диабет связан с несколькими осложнениями, включая нефропатию, ретинопатию, невропатию и атеросклероз [2]. Приблизительно от 60% до 70% всех людей с диабетом в конечном итоге разовьется периферическая невропатия [3]. Растущие эпидемиологические данные свидетельствуют о том, что диабетическая нейропатия и СД2 могут быть связаны с повышенным риском болезни Альцгеймера (БА) [4]. AD — это прогрессирующее заболевание мозга, которое постепенно ухудшает память и способность человека учиться, общаться и выполнять повседневные действия [5].По оценкам, в 2018 году 5,7 миллиона американцев жили с БА [6]. Учитывая высокую распространенность и огромное социальное и экономическое бремя, крайне важно определить эффективный, безопасный и недорогой подход, чтобы отсрочить прогрессирование или предотвратить симптомы этих заболеваний. Однако существующие противодиабетические препараты обладают различными побочными эффектами, и в настоящее время не найдено лечения, предотвращающего или обращающего вспять прогрессирование БА [7, 8]. Учитывая биохимическую связь между AD и T2DM [9, 10], возможно, что существует общая терапевтическая цель для AD и T2DM.Природные биоактивные соединения могут быть альтернативным лечением диабета и новым многообещающим методом лечения БА благодаря их эффективности, меньшему количеству побочных эффектов и легкой доступности [11]. Исследования показали, что благотворное влияние биологически активных соединений может быть обусловлено различными свойствами, такими как антиоксидантное, противовоспалительное и антиапоптотическое действие [11, 12]. Здесь мы рассматриваем множественные полезные эффекты биологически активных соединений и лежащие в их основе механизмы действия в клеточных культурах и на моделях животных o
.
Клеточные и молекулярные механизмы воздействия половых гормонов на нервную систему
1. Введение
Половые гормоны синтезируются из холестерина в основном в гонадах и коре надпочечников. В головном мозге различные половые стероиды также могут далее метаболизироваться до различных нейростероидов или вырабатываться заново в нейронах и глиальных клетках с еще более сильным действием на нервную систему. Механизмы действия половых стероидных гормонов на мозг связаны как с классическими, внутриклеточно-опосредованными эффектами, так и с неклассическими (негеномными) мембранными эффектами из-за их связывания с мембранными рецепторами.Некоторые стероиды способны вызывать быстрые нейротрансмиттерные эффекты. Половые стероиды оказывают разнообразное плейотропное действие на нервную систему: они модулируют основные системы нейромедиаторов, способствуют жизнеспособности нейронов, играют важную роль в миелинизации и влияют на когнитивные процессы. Эстрадиол увеличивает выживаемость и восстановление нейронов. Он защищает нейроны от эксайтотоксического повреждения, токсичности амилоида β (Aβ), окислительного стресса и депривации глюкозы. Защита, вызванная эстрогенами, опосредуется сложными механизмами.Также было обнаружено, что прогестины обладают нейропротекторным действием, аналогичным действию эстрогенов. Андрогены проявляют широкий спектр нейропротекторных эффектов на мотонейроны, включая поддержку выживания клеток, регенерацию аксонов и поддержание дендритов. Связь между половыми стероидами и нейротрофическим фактором головного мозга (BDNF) вызывает растущий интерес из-за роли, которую BDNF играет в патогенезе нейродегенеративных заболеваний.
2. Стероидогенез
Половые гормоны — это стероидные соединения, синтезируемые из холестерина в основном в семенниках, яичниках и коре надпочечников.Мужские половые гормоны (андрогены) и женские половые гормоны (эстрогены и гестагены) имеют общий биосинтетический путь (рис. 1).
Рисунок 1.
Путь биосинтеза половых стероидов. Ферменты показаны следующим образом: (1) фермент расщепления боковой цепи Р450; (2) 3β-гидроксистероиддегидрогеназа; (3) 17β-гидроксистероиддегидрогеназа; (4) 5α-редуктаза; (5) ароматаза. Пунктирная стрелка указывает на плохой поток. Включены не все промежуточные стероиды, метаболические пути и ферменты (с изменениями из Refs.[1, 55]).
Конечный продукт стероидогенеза половых гормонов зависит от того, доступны ли специфические метаболизирующие ферменты в соответствующей клетке [1]. Половые стероиды в крови человека включают андрогены (тестостерон, дегидроэпиандростерон (ДГЭА), дегидроэпиандростерон сульфат (ДГЭАС), андростендион и дигидротестостерон), эстрогены (эстрадиол, эстриол и эстрон) и гестагены (прогестерон и 17α-гидроксиапрогестерон). Основной мужской гормон, тестостерон, вырабатывается клетками Лейдига в яичках.Дигидротестостерон (ДГТ) является мощным андрогеном, полученным из тестостерона с помощью фермента 5α-редуктазы (тип 1 и тип 2) в некоторых периферических тканях, опосредуя некоторые эффекты, вызванные тестостероном. Этот фермент экспрессируется в коже, волосистой части головы, простате, придатке яичка, печени и нервной системе (неокортекс, подкорковое белое вещество и ткани гиппокампа) [2]. DHEA, DHEAS и андростендион секретируются в основном корой надпочечников в одинаковых количествах у обоих полов. DHEA и андростендион — стероиды, участвующие в пути биосинтеза половых гормонов; оба являются первичными эндогенными предшественниками тестостерона и эстрогенов.Хотя они являются слабыми андрогенами, они представляют собой циркулирующие стероиды, которые могут превращаться в активные андрогены и эстрогены в периферических тканях [1, 3].
Эстрогены производятся путем ароматизации андрогенов, в том числе получаемых в результате стероидогенеза надпочечников. Хотя яичники вырабатывают большое количество андрогенов, они секретируют их в кровь мало, а остальная часть ароматизируется до эстрадиола, который является основным эстрогеном. Клетки теки в яичниках синтезируют тестостерон и андростендион, которые затем диффундируют в клетки гранулезы фолликулов.Андростендион превращается в тестостерон, который, в свою очередь, ароматизируется в эстрадиол, попадающий в кровоток. Часть андростендиона ароматизируется до эстрона, который, в свою очередь, превращается в эстрадиол. Ароматизация андрогенов осуществляется под действием фермента ароматазы, который экспрессируется в стероидогенных тканях, головном мозге и нестероидогенных тканях, особенно в жировой и костной. Прогестерон является основным прогестагеном и продуцируется как в клетках теки, так и в гранулезных клетках, надпочечниках и семенниках [1, 3].
Нейроактивные стероиды — это эндогенные нейромодуляторы, синтезируемые мозгом, которые быстро изменяют возбудимость нейронов. Некоторые из них достигают головного мозга через надпочечники и гонады и далее метаболизируются локально, подобно ароматизации тестостерона в эстрадиол [4]. Их называют нейростероидами, поскольку они могут быть получены заново из холестерина в нейронах и глиальных клетках [5]. Синтез нейроактивных стероидов требует транслокации холестерина через митохондриальную мембрану [6].Этот процесс происходит через молекулярный комплекс, образованный транслокаторным белком 18 кДа (TSPO), стероидогенным острым регуляторным белком (StAR), потенциал-зависимым белком анионного канала (VDAC) и белком-переносчиком адениновых нуклеотидов (ANT).
В митохондриях холестерин превращается в прегненолон ферментом расщепления боковой цепи P450 (P450scc). Растворимый прегненолон диффундирует в цитозоль (эндоплазматический ретикулум), где он далее метаболизируется в различные нейроактивные стероиды, такие как прогестерон, 5α-дигидропрогестерон, ДГЭА, андростендион и т. Д.В мозге обнаружен фермент 3β-гидроксистероиддегидрогеназа, необходимый для дальнейшего превращения прегненолона в прогестерон [7]. Фермент 17β-гидроксистероиддегидрогеназа типа 10 катализирует окисление нейроактивных стероидов в митохондриях с NAD + в качестве кофермента. Этот фермент наиболее эффективно катализирует окисление аллопрегнанолона и аллотетрагидродезоксикортикостерона, что важно для гомеостаза этих нейроактивных стероидов [8].
Хотя TSPO высоко экспрессируется в микроглии и астроцитах и менее распространен в нейронах, нейростероидогенез происходит в основном в основных нейронах нескольких областей мозга, которые имеют необходимый набор ферментов для преобразования холестерина в нейроактивные стероиды [9].
3. Механизмы действия
Первое, что делает гормон, — это связывается со специфическими рецепторами на клетке-мишени. Клетки без рецепторов гормона не реагируют на это действие. Рецепторы некоторых гормонов локализуются на клеточной мембране, а другие — в цитоплазме или ядре. После связывания со специфическим рецептором гормон запускает каскад клеточных ответов, которые становятся все более мощными с каждой последующей стадией. Таким образом, даже небольшие концентрации гормона могут дать значительный эффект [3].
3.1. Геномное действие через стероидные рецепторы
Согласно классической геномной теории действия, половые гормоны как стероидные гормоны связываются преимущественно со специфическими белковыми рецепторами внутри клетки, а не с рецепторами, расположенными на клеточной мембране. Эти гормоны жирорастворимы и могут легко проходить через клеточную мембрану и связываться со специфическими рецепторами в цитоплазме. Однако в зависимости от стероида и ткани несвязанные стероидные рецепторы также могут быть расположены в ядре.Конкретное распределение рецептора между цитоплазмой и ядром варьируется. Когда цитоплазматические рецепторы связываются со своими специфическими лигандами стероидных гормонов, они перемещаются в ядро. В зависимости от механизма действия и субклеточного распределения ядерные рецепторы можно разделить как минимум на две группы [10]. Ядерные рецепторы, которые связываются со стероидными гормонами, классифицируются как рецепторы типа I. Только рецепторы типа I имеют белок теплового шока (HSP), связанный с неактивным рецептором, который высвобождается при взаимодействии рецептора с лигандом.Ядерные рецепторы II типа не имеют HSP и, в отличие от классических рецепторов I типа, расположены в ядре клетки. Активированные белковые комплексы гормон-рецептор затем связываются со специфическим регуляторным участком ДНК, называемым гормональным ответным элементом, путем активации или ингибирования транскрипции определенных генов и образования информационной РНК. Позже, после продолжительного периода времени (обычно от нескольких часов до нескольких дней), отсчитываемого от попадания гормона в клетку, в клетке развиваются новые белки, которые изменяют функции клетки.
Сложность действия стероидов может быть объяснена обилием идентифицированных стероидных рецепторов и их сродством к гормону. Избыток / дефицит соответствующего полового стероида регулирует количество активных рецепторов (понижающая регуляция / повышающая регуляция) в клетках-мишенях. Тестостерон и ДГТ осуществляют свои функции посредством связывания с рецептором андрогена (AR), что приводит к конформационному изменению рецептора и перемещению комплекса андроген / AR из цитозоля в ядро.Различные корегуляторы AR могут дополнительно модулировать регуляцию транскрипции генов-мишеней [11]. Рецепторы AR экспрессируются в нейронах и глиальных клетках, и их экспрессия может регулироваться повреждением и концентрацией циркулирующего тестостерона [12, 13, 14]. Уровень мРНК AR снижается после орхидэктомии и после аксотомии [12]. Уровни АР также снижаются с возрастом, особенно в базальном ядре Мейнерта (которое дегенерирует при болезни Альцгеймера (БА)) и диагональной полосе Брока [15].
Рецептор эстрогена-α (ERα) был охарактеризован как внутриклеточный, регулируемый лигандом фактор транскрипции, расположенный преимущественно в ядре [16].Было показано, что после связывания с эстрадиолом димеры ERα регулируют экспрессию генов посредством взаимодействия с элементами ответа на эстроген. После серии открытий был идентифицирован структурно родственный рецептор эстрогена-β (ERβ) [17]. Участки экспрессии рецепторов эстрогена, идентифицированные в головном мозге, включали, среди прочего, гипоталамус, гипофиз и преоптическую область, которые, как было известно на основе серии исследований повреждений и стимуляции, влияют на физиологию и поведение, связанное с эндокринной функцией [18].Помимо большого количества различных изоформ, классические внутриклеточные рецепторы также имеют множество вариантов сплайсинга, которые были изучены и охарактеризованы. Например, для эстрогенов, помимо изоформ ERα и ERβ, несколько вариантов сплайсинга (например, ERαΔ4) могут инициировать передачу сигналов от мембраны [19]. Эксперименты показали, что один и тот же белок может опосредовать как внутриклеточное, так и мембранное действие эстрадиола. Для прогестерона был идентифицирован целый новый класс рецепторов прогестерона (PR) — мембранные PR, локализованные на мембране и участвующие в репродуктивной деятельности прогестерона [20].
3.2. Негеномное действие
Классический геномный механизм действия только стероидных гормонов не может объяснить все последующие изменения в клетках-мишенях; следовательно, он был обновлен, чтобы включить дополнительное (не классическое) объяснение быстрого негеномного, инициируемого мембраной действия. На протяжении десятилетий известно, что стероидные гормоны вызывают резкие изменения (в течение нескольких минут) физиологических функций [21], нейрональной активности [22] и поведения [23].
Недавнее исследование продемонстрировало, что стероиды могут действовать «подобно нейротрансмиттерам», синтезируются в точных пространственных точках в нервных цепях мозга и действуют в течение нескольких минут как локальные нейромодуляторы, которые быстро регулируют когнитивные функции и поведение [24, 25, 26 , 27].
Некоторые стероиды, такие как прогестерон, способны вызывать быстрые нейротрансмиттерные эффекты, подобные эффектам дофамина или глутамата, которые изменяют активность нейронных систем через несколько типов рецепторов [19, 25, 28]. Некоторые из этих стероидных рецепторов были классифицированы как внеядерные или мембранные рецепторы, которые передают сигнал через G-белки или другие системы вторичных мессенджеров [29, 30]. Недавно появились доказательства того, что эти классические стероидные рецепторы связываются с элементами ответа на ДНК, чтобы регулировать экспрессию генов, показывая, что они содержат последовательности пальмитоилирования, позволяющие им перемещаться к плазматической мембране для быстрого изменения клеточной активности [19, 31].После передачи эти ядерные факторы транскрипции взаимодействуют с другими белками, чтобы инициировать их передачу сигналов на уровне плазматической мембраны. Отсюда внутриклеточные сигнальные каскады с участием эффекторов (например, митоген-активируемой протеинкиназы (MAPK) и белка, связывающего элемент ответа цАМФ (CREB)) инициируются через трансактивацию рецепторов, связанных с поверхностью клетки, в первую очередь метаботропных рецепторов глутамата (mGluRs ). Впоследствии эстроген-инициируемая мембрана передача сигналов может, в свою очередь, активировать регуляторный участок ДНК и запускать процессы транскрипции.
Современное понимание клеточного ответа на действие стероидов состоит в том, что он происходит в те же временные рамки, что и рецепторы, связанные с G-белком, влияющие на различные клеточные функции, такие как закрытие мембранных каналов, увеличение высвобождения внутриклеточного кальция, активация тирозин-протеинкиназа (Src), MAPK и другие [27]. Многие исследования поддерживают модель интегрированной передачи сигналов, которая связывает каскады передачи сигналов с транскрипцией в ядре, обеспечивая комплексное представление о передаче сигналов гормонов в головном мозге [32].
В последнее время обширные исследования, посвященные быстрому негеномному действию эстрогенов, подняли вопрос о том, насколько быстро может происходить повышение уровня этих стероидов в мозге. Конечно, эстрогены, как и любые другие стероиды, не могут храниться в синаптических пузырьках до их быстрого высвобождения из-за их липофильной природы [4]. Поэтому было высказано предположение, что быстрое действие эстрогенов требует соответствующего быстрого изменения локальной концентрации стероидов за счет быстрых изменений скорости их синтеза путем превращения андрогенов [24, 33], что подразумевает изменения в активности ароматазы.Изменения активности ароматазы отражают изменения концентрации белка ароматазы. Например, половые стероиды контролируют экспрессию гипоталамической ароматазы у большинства позвоночных: у кастрированных самцов животных обнаруживается слабая экспрессия ароматазы, тогда как замещение тестостерона значительно увеличивает активность белка ароматазы и ферментов [34, 35]. Существуют убедительные доказательства того, что активность ароматазы может быть быстро модулирована посредством трансляционных модификаций, в первую очередь посредством фосфорилирования. Быстрая модуляция активности ароматазы путем фосфорилирования является широко распространенным механизмом, присутствующим в некоторых тканях различных видов, включая человека [4].Ферментативные изменения приводят к быстрой местной модуляции доступности эстрогена и, следовательно, к модификации клеточных эстроген-зависимых процессов, которые не опосредуются геномными действиями этих стероидов. Процессы фосфорилирования / дефосфорилирования обеспечивают новый широко распространенный механизм, с помощью которого концентрация эстрогена может быстро изменяться в мозге и других тканях.
Хотя большинство исследований нейротрансмиттероподобного действия стероидных гормонов сосредоточено на половых гормонах и репродуктивной функции, другие стероиды также вызывают эффекты посредством неклассических механизмов.Как и эстрогены и прогестины, глюкокортикоиды могут воздействовать на мембрану, изменяя физиологию, действуя больше как нейротрансмиттеры, чем классические стероидные гормоны.
Нейростероиды также способны взаимодействовать с рецепторами нейротрансмиттеров клеточной поверхности, чтобы модулировать физиологию нервных клеток. Два эндогенных нейростероида, прегненолона сульфат и прегнанолон сульфат, могут усиливать или ингибировать реакции рецепторов N-метил-D-аспартата (NMDA) [36]. GABA A рецепторы представляют собой одну из наиболее сложных рецепторных структур нейротрансмиттеров, несущих множественные сайты связывания для аллостерических модуляторов, нейроактивных соединений и нейроактивных стероидов [37].Было показано, что аллопрегнанолон способствует нейрогенезу как в нейропрогениторных клетках грызунов, так и в человеческих клетках-предшественниках, наиболее вероятно за счет связывания с рецептором GABA A [38]. Модуляция активности рецепторов нейрохимическими веществами, такими как аллопрегнанолон, широко изучалась в контексте нейродегенеративных расстройств [39].
Другой механизм действия стероидов действует на уровне микротрубочек через предложенный рецепторный белок типа 2 (MAP2), связанный с микротрубочками [40].Нейрональные микротрубочки играют важную роль в росте и поддержании нейритов во время дифференцировки нейронов. Они состоят из тубулина и белков, связанных с микротрубочками (MAP). MAP определяют форму нейронов и контролируют баланс между жесткостью и пластичностью нейронных процессов. Нейростероиды могут участвовать в образовании и стабилизации микротрубочек и, следовательно, в пластичности и функции нейронов [40]. Экспериментальные данные демонстрируют, что лечение прогестероном уменьшало вызванную повреждением потерю MAP2 [41].
4. Биологические эффекты половых гормонов на нервную систему
Тестостерон и его метаболит эстрадиол вызывают многочисленные эффекты в критические периоды пре- и перинатального развития мозга (организационные эффекты), которые необходимы для половой дифференциации мозга. Воздействие тестостерона является важным условием маскулинизации мозга. Объем ядра, морфология нейронов и сложность астроцитов являются примерами широкого спектра эффектов, с помощью которых тестостерон и эстрадиол могут вызывать необратимые изменения в функции нейронов [42].У развивающегося самца крысы тестостерон, секретируемый семенниками, не связывается α-фетопротеином и свободно проникает в мозг, где он локально превращается в эстрадиол в определенных ядрах. Следовательно, у новорожденных мужчин уровень эстрадиола более чем в два раза выше, чем у женщин, в областях мозга, подверженных половому признаку [43]. Высокие уровни ER сосредоточены в одних и тех же областях мозга, и ER необходим для передачи стероидного сигнала [44]. Прирост или потеря функции при воздействии эстрадиола в процессе развития соответствует специфическим клеточным морфологическим изменениям, наблюдаемым в критический период, а дендритные шипы и астроциты, наблюдаемые в каждой области мозга, сохраняют эту «память» о раннем воздействии стероидов [42].
4.1. Эффекты женских половых стероидов
Принято считать, что эстроген действует как условный нейропротектор со сложным паттерном биологических действий, которые модулируются несколькими взаимодействующими факторами [45]. Было обнаружено, что введение эстрадиола увеличивает выживаемость и восстановление нейронов у взрослых животных и различных моделей поражения [46, 47]. Эстрадиол защищает нейроны от эксайтотоксического повреждения при судорогах и инсульте, а также при БА [48]. Один из предполагаемых механизмов этого эффекта — способность эстрогенов усиливать экспрессию и высвобождение нейропептида Y (NPY), поскольку NPY обладает противовозбуждающим действием [49].Было обнаружено, что in vitro эстрадиол защищает нейроны от токсичности глутамата и пептида Aβ, окислительного стресса и депривации глюкозы [50, 51, 52, 53]. Состояние защиты, индуцированное эстрогеном, опосредуется сложными механизмами, которые сходятся при регуляции функции митохондрий. Эстроген сохраняет уровни АТФ за счет повышенного окислительного фосфорилирования и снижения активности АТФазы, тем самым увеличивая эффективность митохондриального дыхания. Эстроген увеличивает антиапоптотические белки, Bcl-2 и Bcl-xL, которые предотвращают образование пор перехода проницаемости, защищая от вызванного эстрогеном увеличения секвестрации митохондриального Ca 2+ и запуска апоптотических процессов [54].Следовательно, пониженный уровень эстрогена, скорее всего, может способствовать повышенному риску развития нейродегенеративных заболеваний, особенно у женщин в постменопаузе [52, 55].
Предполагается, что в дополнение к прямому действию на нейроны эстрогены могут влиять на астроциты, стимулируя их высвобождение защитных факторов роста и регулируя гены астроцитов и белки, связанные с контролем уровня глутамата. Другие механизмы, задействованные здесь, могут включать противовоспалительный эффект, связанный с подавлением микроглии, воспалительных цитокинов и выработки свободных радикалов, которые вызывают воспалительное повреждение нейронов, эффекты на эндотелиальные клетки, реализуемые за счет увеличения митохондриальной эффективности и стимуляции ангиогенеза, влияние генома на гены антиапоптотических белков семейства Bcl и снижение апоптотических тенденций и эффекта улавливания свободных радикалов.Это гипотетические модели нейропротекции эстрогеном при церебральной ишемии и других нейродегенеративных расстройствах, таких как болезнь Паркинсона (БП) и БА [52, 56].
Появляется все больше доказательств того, что эстроген может иметь нейропротекторную роль при БП. Экспериментальные исследования показали, что эстроген обладает нейропротекторным действием при нигростриатальных поражениях, вызванных 1-метил-4-фенил-1,2,3,6-тетрагидропиридином (МРТР), на животной модели идиопатической БП [57, 58]. В этих и других исследованиях использовался 17β-эстрадиол, и было показано, что его действие стереоспецифично.Изомер со слабой эстрогенной активностью, 17α-эстрадиол, был неэффективен в отношении предотвращения вызванной МРТР потери дофамина [52]. Следует отметить, что рецепторы ERα и ERβ редко локализованы в полосатом теле и черной субстанции мышей, и лечение МРТР или эстрогенами не меняет распределения и плотности рецептора эстрогена. Несмотря на низкую доступность ER в этих частях мозга, эстрогену удалось вызвать защитное действие на полосатое тело против вызванной МРТР потери дофаминергических нейронов [59].
Исследования на людях показали, что краткосрочная терапия эстрогенами у женщин в постменопаузе увеличивает доступность переносчиков дофамина в хвостатой скорлупе [60] и что у женщин, которые принимали заместительную терапию эстрогенами в постменопаузе, вероятность развития БП была ниже, чем у тех, кто этого не делал [61] ,
Имеются данные об индукции дифференцировки нервных стволовых клеток человека, которые развиваются в нейронах тирозингидроксилазы (дофаминергических), и эффект блокируется применением антагониста рецептора эстрогена [62, 63].
Поскольку предполагается, что окислительный стресс играет важную роль в процессах дегенерации нейронов при БП, интересно, что эстрогены подавляют продукцию свободных радикалов и защищают нейроны полосатого тела от окислительного стресса, обеспечивая другой механизм нейрозащиты эстрогена при БП [64 , 65].
Недавние исследования как на животных, так и на людях предоставили дополнительные доказательства, подтверждающие потенциально полезную защитную роль эстрогена при БА. Механизмы защиты эстрогенов при БА не ясны.Было показано, что на молекулярном уровне эстроген усиливает активацию факторов выживания, протеинкиназы B, BDNF [66, 67], вызывая фосфорилирование и дезактивацию киназы гликогенсинтазы (GSK3B) и связанного с Bcl-2 агониста клеточной гибели ( BAD), участвующих в сигнальных путях гибели нейронов [67, 68].
Было также обнаружено, что прогестины обладают нейропротекторным действием, аналогичным действию эстрогенов. Прогестерон стимулирует неврологическое и функциональное восстановление после травм спинного и головного мозга [56, 69] и оказывает нейропротекторное действие при церебральной ишемии [70, 71].
4.2. Действие андрогенов
Действие андрогенов на нервную систему изучено гораздо меньше, чем действие эстрогенов и прогестинов. Андрогены также проявляют широкий спектр нейропротекторных эффектов на мотонейроны, включая поддержание выживания клеток, регенерацию аксонов и поддержание дендритов [72]. Тестостерон влияет на нейропластические изменения в ядрах лимбической системы, особенно в миндалине, ядре ложа терминальной полоски и гиппокампе [73, 74]; он оказывает нейропротекторное действие, стимулируя выживание и регенерацию нейронов после повреждения нерва за счет действий, опосредованных рецептором андрогенов [75, 76].Было обнаружено, что он оказывает защитное действие на апоптоз в клеточных культурах нейронов человека. Этот эффект опосредуется непосредственно рецепторами андрогенов, без ароматизации тестостерона до эстрадиола [77]. Замена тестостерона у гонадэктомированных взрослых мышей-самцов обращает патологические изменения в морфологии позвоночника пирамидных нейронов СА1 гиппокампа. Дендритные шипы специализированы для приема синаптических входов, и изменение морфологии шипов коррелирует с силой и зрелостью каждого синапса [78].Аналогичные данные были получены при экспериментальном повреждении мотонейронов с использованием DHT, снижающего атрофию прилегающих дендритов [79]. Недавние открытия предполагают, что одним из механизмов нейропротективного действия физических тренировок является повышенная продукция DHT в гиппокампе, что свидетельствует об андрогенном опосредовании нейрогенеза рецепторами андрогенов [80].
Андрогены могут регулировать продукцию и уровни Aβ с помощью классического геномного механизма и быстрой негеномной передачи сигналов или посредством ароматизации до эстрадиола и активации эстрогеновых путей [81, 82].Тестостерон может снизить токсичность Aβ в культивируемых нейронах гиппокампа посредством быстрого, независимого от эстрогена механизма [83]. DHT увеличивает Aβ-катаболизирующий фермент неприлизин в культивируемых нейронах посредством AR-зависимого механизма, который способствует деградации Aβ, тем самым снижая уровни Aβ при AD [84].
4.3. Влияние предшественников стероидов
Предшественники эстрогенов, прогестинов и андрогенов (прегненолон и ДГЭА) также влияют на функции нейронов. При введении in vivo прегненолон снижает гистопатологические изменения, защищает нервные ткани от вторичных повреждений и способствует восстановлению двигательных функций после повреждения спинного мозга [85, 86].DHEA — один из первых нейростероидов, обнаруженных в мозге крыс. Документально подтверждены нейропротективные эффекты, вызванные DHEA и его сульфатом DHEAS, определяемые как первичные по их биологическому действию [87]. Оба стероида вносят вклад в дифференцировку и выживание нейронов в культурах клеток [88]; обладают защитным действием на нейроны гиппокампа от токсического действия глутамата [89]; стимулировать рост невритов корковых нейронов эмбрионального мозга крысы [90]; влияют на апоптоз, синтез и секрецию катехоламинов; и обладают антиоксидантным, противовоспалительным и антиглюкокортикоидным действием [87].
Исследования показывают, что это разные механизмы эффектов DHEA и DHEAS. Предполагается, что DHEAS опосредует свои эффекты через рецепторы GABA A , вероятно, путем метаболизма DHEAS в агонист рецептора GABA A , такой как андростерон или андростандиол [91]. Нейропротекторный эффект DHEAS по отношению к цитотоксичности, вызванной рецептором NMDA, вероятно, опосредуется рецептором σ1, в то время как DHEA ингибирует индуцированное NMDA продукцию оксида азота (NO) и активность NO-синтазы рецептором NMDA, модулируя путь передачи сигналов кальция / NO [92].Концентрации DHEA и его сульфата также важны с точки зрения конечного эффекта. Низкие концентрации этих стероидов могут быть нейропротективными, в то время как высокие концентрации DHEA неэффективны или нейротоксичны и приводят к ингибированию комплекса I дыхательной цепи митохондрий [93].
4.4. Взаимодействие между стероидами и нейротрофинами
Недавно исследователи изучали взаимосвязь между стероидами гонад, стероидами надпочечников и BDNF, уделяя особое внимание интерсексуальным различиям и частоте психических заболеваний [94].BDNF принадлежит к семейству нейротрофинов и играет важную роль в выживании, дифференцировке и росте отдельных периферических и центральных нейронов во время развития. BDNF значительно влияет на выживаемость нейронов, воздействуя на мозг взрослого человека через различные типы клеток, включая нейроны, астроциты, олигодендроциты, микроглию и эндотелиальные клетки. Это важно для процесса обучения и улучшения когнитивных функций посредством активации рецептора TrkB [95]. Наши предыдущие данные показали, что отрицательное действие противосудорожного лакозамида на процессы обучения и памяти связано с подавлением экспрессии рецепторной системы лиганда BDNF / TrkB в гиппокампе крыс [96].Половые стероидные гормоны и нейротрофические факторы участвуют в нейроэндокринном контроле репродукции, а также в адаптации мозга во время репродуктивного старения. Существует множество доказательств, показывающих роль BDNF в патогенезе нейродегенеративных заболеваний. Низкие посмертные уровни BDNF в париетальной коре головного мозга были обнаружены у пациентов с легкими когнитивными нарушениями [97] и БА [97, 98].
Исследования показывают, что уровни экспрессии мРНК BDNF и белка в когнитивных областях мозга зависят от региона при назначении заместительной гормональной терапии.Сообщалось, что уровни мРНК BDNF значительно снижены почти во всех слоях гиппокампа и в коре головного мозга у 28-недельных овариэктомированных крыс [99]. Заместительная терапия эстрадиолом обращает этот эффект в гиппокампе, указывая на региональную дивергенцию потребностей яичников в стероидах для экспрессии BDNF. После гонадэктомии уровни мРНК BDNF значительно снижаются на 7-й постнатальный день у крысят-самцов, но после лечения бензоатом эстрадиола уровни были аналогичны уровням у интактных животных.Авторы продемонстрировали, что ERα и BDNF локализованы в одних и тех же клетках (пирамидных клетках субрегиона CA3 и в меньшей степени в CA1) в развивающемся гиппокампе [100].
Эстрогены участвуют в повышении уровня мРНК и белка BDNF в гиппокампе у тренирующихся животных. Влияние упражнений на активацию BDNF уменьшилось после 7 недель лишения эстрогена. Упражнения в сочетании с длительной заменой эстрогена увеличивают белок BDNF по сравнению с эффектами замены только эстрогена [101].
Андрогены также влияют на экспрессию BDNF; некоторые из их эффектов на нервную систему, скорее всего, будут реализованы через влияние на выработку этого нейротрофина. Показано, что введение тестостерона увеличивает уровень белка BDNF в мотонейронах спинномозгового ядра луковично-кавернозного канала кастрированных самцов крыс [102]. Гонадэктомия вызывает значительное снижение уровней белка BDNF и его нижележащего целевого белка постсинаптической плотности 95 (PSD-95) в области CA1 гиппокампа, что устраняется заменой тестостерона [78].Знание взаимодействий между BDNF и половыми стероидами может иметь важное значение для понимания роли BDNF в развитии мозга, адаптации во время старения и патогенезе нейродегенеративных заболеваний.
5. Заключение
Функции половых гормонов превышают пределы воспроизводства, поскольку они регулируют жизненно важные нейрональные и глиальные функции. Хронические эффекты нейростероидов обусловлены как геномными (классические внутриклеточные стероидные рецепторы), так и негеномными быстрыми эффектами (ионные каналы и мембранные рецепторы) в головном мозге.
Некоторые из гипотетических моделей нейропротекции эстрогеном включают сложные механизмы, которые сходятся на регуляции функции митохондрий, сохраняя уровни АТФ за счет увеличения окислительного фосфорилирования и увеличения антиапоптотических белков семейства Bcl. Эстроген стимулирует астроциты к высвобождению защитных факторов роста и оказывает противовоспалительное действие, связанное с подавлением микроглии и воспалительных цитокинов. Он подавляет выработку свободных радикалов и защищает нейроны полосатого тела от окислительного стресса, обеспечивая еще один механизм нейрозащиты при БП.Женские половые стероиды способствуют выживанию клеток за счет активации протеинкиназы B и активации BDNF; они инактивируют GSK3B и BAD, участвующие в сигнальных путях гибели нейронов при БА. Андрогены также обладают нейропротективным действием на мотонейроны, включая поддержку выживания нейронов, регенерацию аксонов и поддержание дендритов. Тестостерон может ослабить токсичность Aβ и снизить уровень Aβ при AD.
Несмотря на растущее количество исследований половых гормонов и нейростероидов в последние годы и продолжающееся открытие биохимических механизмов действия, их роль в нейродегенеративных процессах остается неопределенной.Дальнейшее выяснение клеточных и молекулярных механизмов, ответственных за влияние нейростероидов на нормальную функцию нейрональных и глиальных клеток, дало бы важную информацию, связанную с разработкой новых терапевтических стратегий, направленных на отсрочку наступления и замедление прогрессирования когнитивных дисфункций и нейродегенеративных заболеваний. ,
.