МЕЖПОЛУШАРНАЯ АСИММЕТРИЯ • Большая российская энциклопедия
В книжной версии
Том 19. Москва, 2011, стр. 575
Скопировать библиографическую ссылку:
Авторы: В. С. Ротенберг
МЕЖПОЛУША́РНАЯ АСИММЕ́ТРИЯ мозга, распределение определённых психич. функций и функций организации поведения между отдельными структурами правого и левого полушарий коры головного мозга человека. Первые представления о М. а. сложились в сер.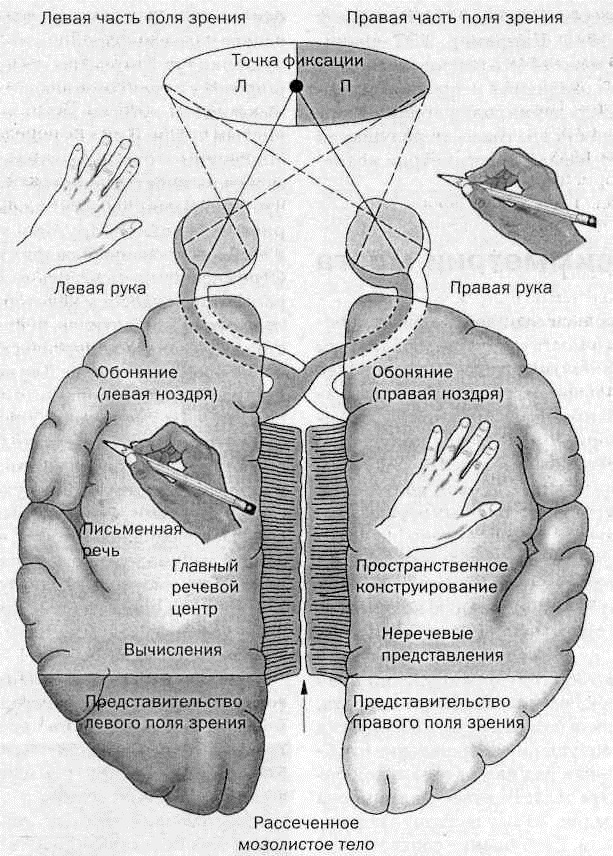 19 в. на основе неврологич. исследований, показавших, что повреждение височной доли левого полушария приводит к нарушению речи и её восприятия, тогда как при повреждении теменных отделов правого полушария нарушается ориентация в пространстве, включая пространство собств. тела. Решающим шагом в развитии представлений о М. а. стали исследования Р. У. Сперри и его сотрудников, которые в целях предотвращения генерализованных эпилептич. припадков провели у нескольких пациентов рассечение межполушарных нервных связей (мозолистого тела и передней спайки) и изучили функции разделённых левого и правого полушарий. Др. методологич. подходом было врем. раздельное выключение полушарий в процессе электросудорожной терапии аффективных расстройств. Ныне исследования М. а. проводятся в осн. на целом мозге здоровых и больных людей с помощью методов томографии и функционального магнитного резонанса.
19 в. на основе неврологич. исследований, показавших, что повреждение височной доли левого полушария приводит к нарушению речи и её восприятия, тогда как при повреждении теменных отделов правого полушария нарушается ориентация в пространстве, включая пространство собств. тела. Решающим шагом в развитии представлений о М. а. стали исследования Р. У. Сперри и его сотрудников, которые в целях предотвращения генерализованных эпилептич. припадков провели у нескольких пациентов рассечение межполушарных нервных связей (мозолистого тела и передней спайки) и изучили функции разделённых левого и правого полушарий. Др. методологич. подходом было врем. раздельное выключение полушарий в процессе электросудорожной терапии аффективных расстройств. Ныне исследования М. а. проводятся в осн. на целом мозге здоровых и больных людей с помощью методов томографии и функционального магнитного резонанса.
Общим свойством структур правого полушария является целостное восприятие реальности как интегрального образа (гештальта) и организация адекватных ему целостных поведенческих реакций, тогда как левое полушарие выделяет и анализирует отд. составные элементы картины мира и организует последовательную целенаправленную деятельность. Так, фонематич., синтаксич. и грамматич. организация речи, побуквенный анализ вербального материала является функцией левой височной доли, а правая обеспечивает восприятие слова как целого. Правая также ответственна за различение интонаций и восприятие мелодий, левая – за восприятие муз. ритмов.
Высокодифференцированные моторные функции контролируются теменной и лобной долями левого полушария, а правая теменная доля отвечает за гармоничные, координированные движения, ориентацию в пространстве. Переднетеменные отделы левого полушария обеспечивают направленное внимание и выделение фигуры из фона, а симметричные им отделы правого полушария ответственны за широту поля внимания. Задние и центральные отделы правого полушария превосходят соответствующие структуры левого в способности к восприятию сложной зрительно-пространственной информации. Правая височная доля ответственна за восприятие и опознание целых объектов, правая теменная – за восприятие их комбинаций, а соответствующие отделы левого полушария – за восприятие их фрагментов. Правое полушарие воспринимает любую информацию быстрее левого, что обеспечивает гибкость и динамизм невербального поведения, особенно в стрессовых ситуациях. Функцией правого полушария являются восприятие и регуляция эмоций.
Переднетеменные отделы левого полушария обеспечивают направленное внимание и выделение фигуры из фона, а симметричные им отделы правого полушария ответственны за широту поля внимания. Задние и центральные отделы правого полушария превосходят соответствующие структуры левого в способности к восприятию сложной зрительно-пространственной информации. Правая височная доля ответственна за восприятие и опознание целых объектов, правая теменная – за восприятие их комбинаций, а соответствующие отделы левого полушария – за восприятие их фрагментов. Правое полушарие воспринимает любую информацию быстрее левого, что обеспечивает гибкость и динамизм невербального поведения, особенно в стрессовых ситуациях. Функцией правого полушария являются восприятие и регуляция эмоций.
Функцией левых фронтоорбитальных (лобных) отделов мозга являются логич.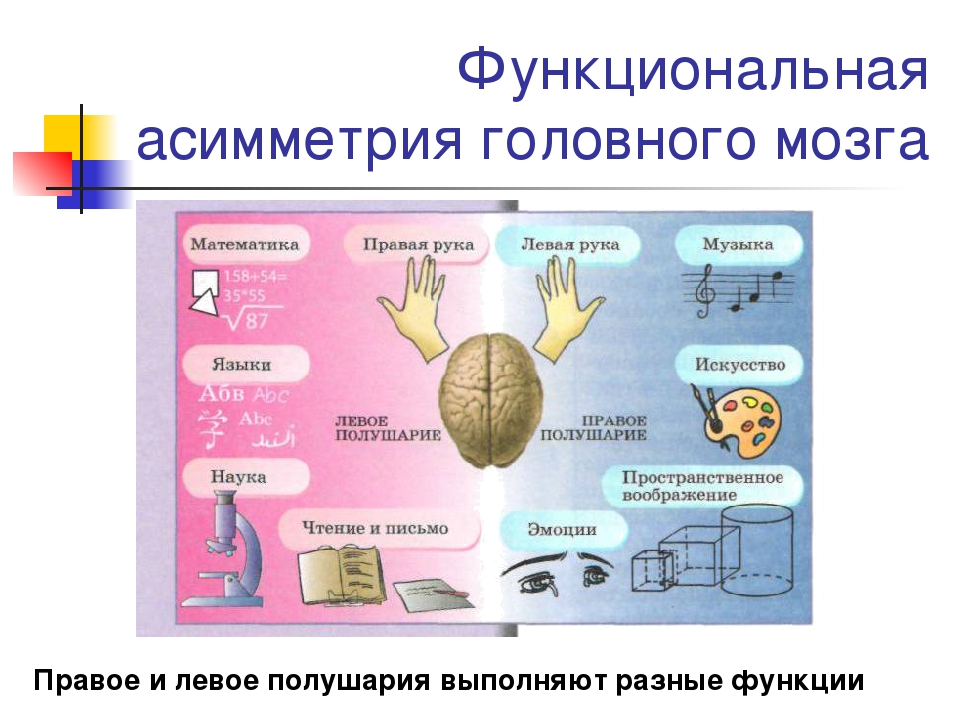 мышление, вероятностный прогноз, обобщения, представления о «стреле времени» от прошлого к будущему, формирование Я-концепции (осознанного представления человека о себе), когнитивный контроль поведения. Правые фронтоорбитальные отделы мозга обеспечивают чувство эмпатии, понимание представлений др. людей (индивидуальные теории психики), метафор, юмора. Левые лобные отделы мозга из всех связей между предметами и явлениями выбирают немногие, позволяющие построить внутренне непротиворечивую модель реальности, однозначно понимаемый контекст, лежащий в основе анализа причинно-следственных отношений и осознанного целенаправленного поведения. Правые лобные отделы охватывают связи между предметами и явлениями в гораздо более полном объёме и создают многозначный контекст, необходимый для полноценной интеграции в мире, межличностных отношений и творчества.
мышление, вероятностный прогноз, обобщения, представления о «стреле времени» от прошлого к будущему, формирование Я-концепции (осознанного представления человека о себе), когнитивный контроль поведения. Правые фронтоорбитальные отделы мозга обеспечивают чувство эмпатии, понимание представлений др. людей (индивидуальные теории психики), метафор, юмора. Левые лобные отделы мозга из всех связей между предметами и явлениями выбирают немногие, позволяющие построить внутренне непротиворечивую модель реальности, однозначно понимаемый контекст, лежащий в основе анализа причинно-следственных отношений и осознанного целенаправленного поведения. Правые лобные отделы охватывают связи между предметами и явлениями в гораздо более полном объёме и создают многозначный контекст, необходимый для полноценной интеграции в мире, межличностных отношений и творчества.
В первые 2 года у ребёнка доминирует правое полушарие, обеспечивая целостную оценку ситуации и эмоциональный контакт с матерью, под влиянием которого формируются связи в правом полушарии. Позднее начинает доминировать левое полушарие, обеспечивающее развитие речи, логич. мышления и рефлексии. Последними, уже в подростковом возрасте, развиваются фронто-орбитальные отделы правого полушария. При раннем повреждении левого полушария правое может взять на себя выполнение вербальных функций, но при поражении правого левое не берёт на себя функцию ориентации в пространстве.
Современные аспекты изучения функциональной межполушарной асимметрии мозга (обзор литературы) Текст научной статьи по специальности «Клиническая медицина»
УДК 612.821
СОВРЕМЕННЫЕ АСПЕКТЫ ИЗУЧЕНИЯ ФУНКЦИОНАЛЬНОЙ МЕЖПОЛУШАРНОЙ АСИММЕТРИИ МОЗГА (ОБЗОР ЛИТЕРАТУРЫ)
© 2016 г. Ю. П. Игнатова, И. И. Макарова, О. Ю. Зенина, А. В. Аксенова
Ю. П. Игнатова, И. И. Макарова, О. Ю. Зенина, А. В. Аксенова
Тверской государственный медицинский университет, г. Тверь
С позиций классических и современных представлений вопрос организации функциональной межполушарной асимметрии вызывает большой интерес у разных специалистов. В настоящей статье представлен обзор современных данных о стационарных и динамических свойствах функциональной специализации полушарий головного мозга. Освещены вопросы функциональной недостаточности межполушарных взаимодействий при некоторых заболеваниях, представлен сравнительный анализ показателей социально-психологической адаптации у лиц с доминированием одного из полушарий, а также особенности латерализации здорового мужского и женского мозга и при патологии. Авторы статьи отмечают ведущую роль в межполушарном взаимодействии мозолистого тела, особенности межполушарной асимметрии правшей и левшей как отражение взаимодействия коры и регуляторных систем мозга во время спокойного бодрствования, активной деятельности и засыпания. В статье обобщены особенности латерального устройства мозга у спортсменов некоторых видов спорта и межполушарные взаимодействия в различные фазы десинхроноза, при старении организма в норме и стрессирующих экологических условиях. Авторы затрагивают вопрос о связи беременности и латерализации головного мозга, а также функциональной специализации полушарий с асимметрией вегетативной нервной системы.
В статье обобщены особенности латерального устройства мозга у спортсменов некоторых видов спорта и межполушарные взаимодействия в различные фазы десинхроноза, при старении организма в норме и стрессирующих экологических условиях. Авторы затрагивают вопрос о связи беременности и латерализации головного мозга, а также функциональной специализации полушарий с асимметрией вегетативной нервной системы.
Ключевые слова: функциональная межполушарная асимметрия, стабильность, динамичность, патология, адаптация
CURRENT ASPECTS OF FUNCTIONAL HEMISPHERIC ASYMMETRY STUDYING
(LITERATURE REVIEW) J. P. Ignatova, 1.1. Makarova, O. J. Zenina, A. V. Aksenova
Tver State Medical University, Tver, Russia
According to the classical and modern concepts the organization of functional hemispheric asymmetry is an interesting issue for different specialists.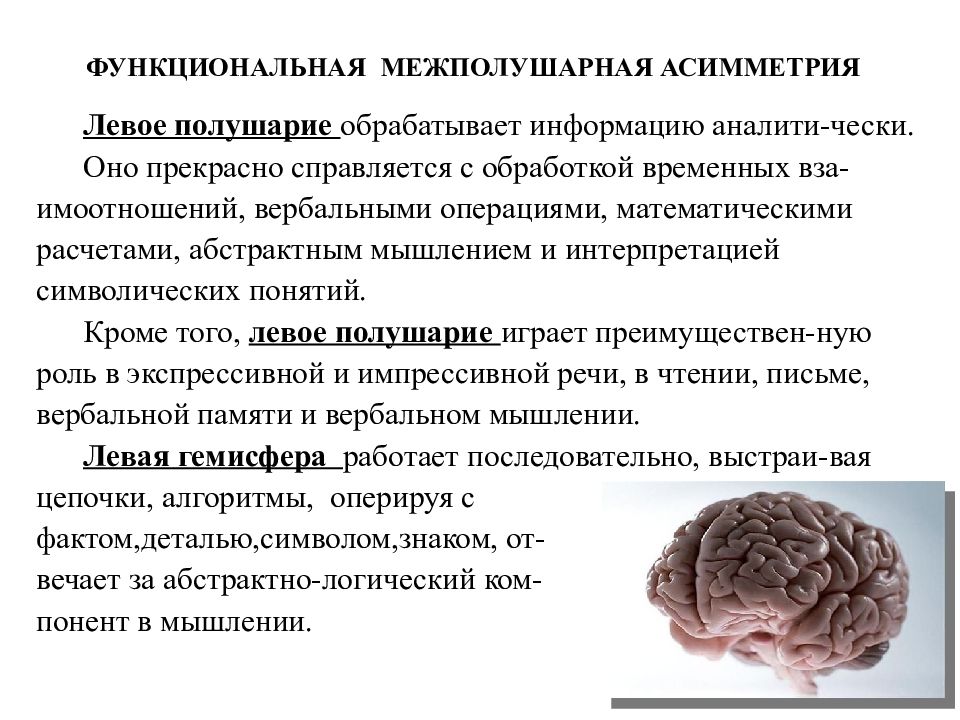 This article reviews current data on fixed and dynamic properties of the cerebral hemispheres functional specialization. The article covers the issues of functional insufficiency of hemispheric interrelations associated with some diseases, gives comparative analysis of parameters of social and psychological adaptation of individuals with one-hemisphere dominance, and presents features of lateralization of brain function in healthy males and females and those with pathology. The authors emphasize the leading role of the corpus collosum in hemispheric interrelations, reveal the features of hemispheric asymmetry of right-handed and left-handed individuals based on interaction of the cortex and the brain regulatory systems during quiet wake, dynamism and falling asleep. The article also focuses on the features of brain function lateralization of some athletes and hemispheric interrelations in different phases of desynchronosis and during organism ageing in normal and stressful ecological conditions.
This article reviews current data on fixed and dynamic properties of the cerebral hemispheres functional specialization. The article covers the issues of functional insufficiency of hemispheric interrelations associated with some diseases, gives comparative analysis of parameters of social and psychological adaptation of individuals with one-hemisphere dominance, and presents features of lateralization of brain function in healthy males and females and those with pathology. The authors emphasize the leading role of the corpus collosum in hemispheric interrelations, reveal the features of hemispheric asymmetry of right-handed and left-handed individuals based on interaction of the cortex and the brain regulatory systems during quiet wake, dynamism and falling asleep. The article also focuses on the features of brain function lateralization of some athletes and hemispheric interrelations in different phases of desynchronosis and during organism ageing in normal and stressful ecological conditions. The authors address the issue of the relationship of pregnancy and the brain lateralization, as well as the functional specialization of the cerebral hemispheres with the asymmetry of the autonomic nervous system.
The authors address the issue of the relationship of pregnancy and the brain lateralization, as well as the functional specialization of the cerebral hemispheres with the asymmetry of the autonomic nervous system.
Key words: functional hemispheric asymmetry, stability, dynamism, pathology, adaptation
Библиографическая ссылка:
Игнатова Ю. П., Макарова И. И., Зенина О. Ю., Аксенова А. В. Современные аспекты изучения функциональной межполушарной асимметрии мозга (обзор литературы) // Экология человека. 2016. № 9. С. 30-39.
Ignatova J. P., Makarova I. I., Zenina O. J., Aksenova A. V. Current Aspects of Functional Hemispheric Asymmetry Studying (Literature Review). Ekologiya cheloveka [Human Ecology]. 2016, 9, pp. 30-39.
Одним из основных принципов функционирования полушарий головного мозга является асимметрия.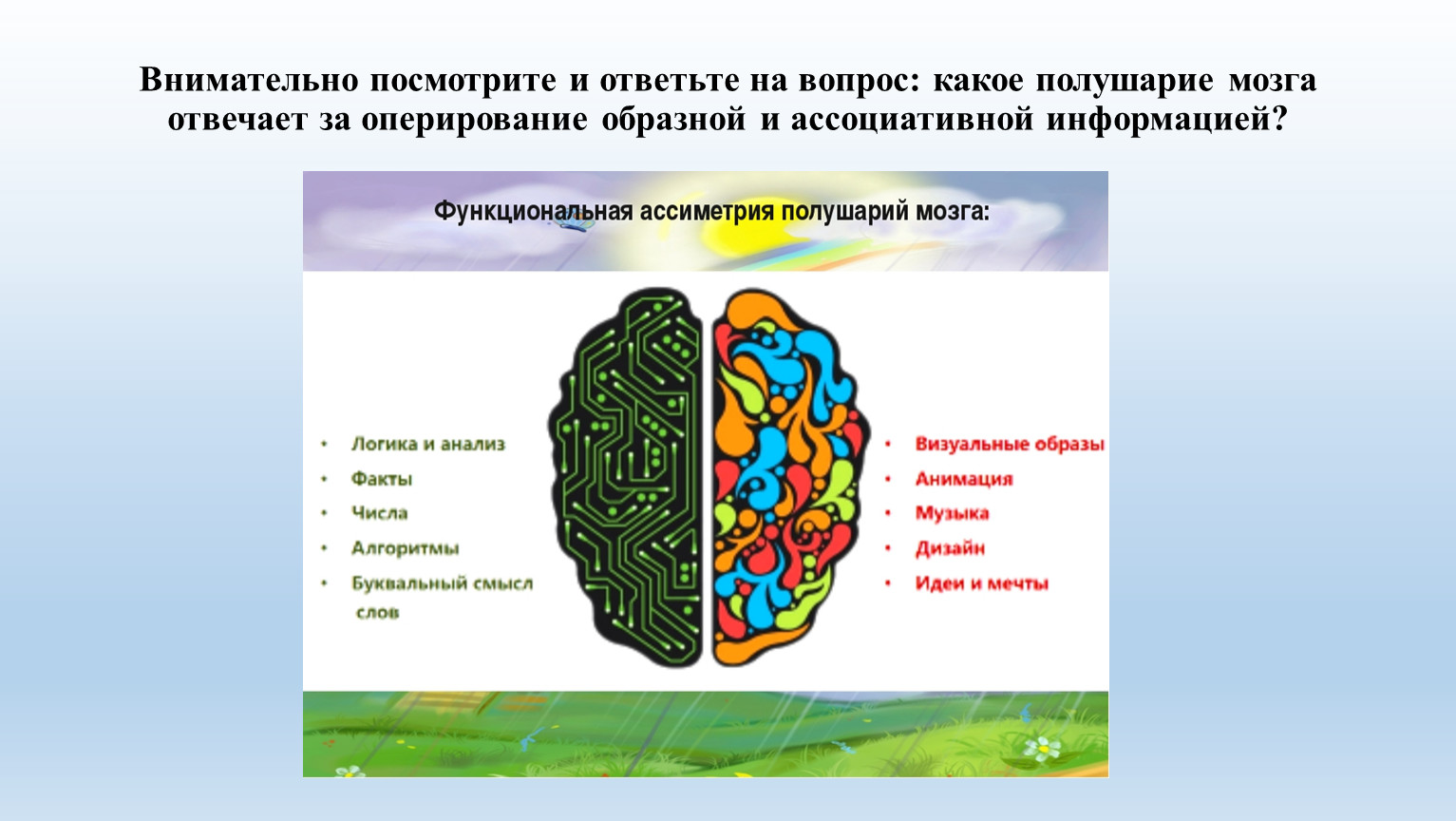 Неравнозначность левой и правой гемисфер мозга человека, сочетаясь с феноменом доминирования одной из них, влияет на специфику построения и восприятия картины мира, характер реагирования и поведения [4].
Неравнозначность левой и правой гемисфер мозга человека, сочетаясь с феноменом доминирования одной из них, влияет на специфику построения и восприятия картины мира, характер реагирования и поведения [4].
Понятие «асимметрия» в переводе с греческого означает неравномерность, разнообразие, разнопо-добие. Первым, кто высказал предположение о том, что все способности человека предопределяются активностью конкретных участков мозга, был немецкий анатом Ф. Галль (1810—1820). Дальнейшее изучение проблемы асимметрии мозга связано с та-
кими именами, как М. Дакс (1836), А. Ваган (1844), Э. Обуртен (1861), П. Брок (1861), Дж. Джексон (1869), К. Вернике (1874). Важные для понимания функционирования полушарий исследования были проведены в 1960-х годах американскими учеными Р. Сперри и М. Газзанигой на больных с расщепленным мозгом. Позднее проблема межполушарной асимметрии стала изучаться прежде всего как проблема функциональной специфичности полушарий. Эти представления построены на нейропсихологической теории мозговой организации высших психических функций, сформулированной А. Р. Лурия.
Эти представления построены на нейропсихологической теории мозговой организации высших психических функций, сформулированной А. Р. Лурия.
Согласно классическим представлениям, передние отделы левого полушария обеспечивают анализ
причинно-следственных связей и целенаправленную деятельность, формирование высоко упорядоченного однозначно понимаемого контекста, в котором связи между элементами не создают внутренних противоречий и который обеспечивает последовательный логический анализ. Общим же свойством всех структур правого полушария является целостность — целостность восприятия и целостность поведения, не подчиняющаяся простым алгоритмам. Правые передние отделы мозга обеспечивают формирование многозначного контекста и тем самым гармоничную интеграцию человека в мире и творчество [24]. Функциональная недостаточность правополушарных механизмов лежит в основе дезадаптации и многих психических и психосоматических заболеваний. Так, в работе Т. В. Черниговской и соавт. [36] по изучению восприятия интонаций русского языка в норме и при заболевании шизофренией отмечено, что у обследуемых пациентов имеется картина сглаженности функциональной асимметрии мозга, в то время как у здоровых испытуемых показано доминирование правого полушария.
Так, в работе Т. В. Черниговской и соавт. [36] по изучению восприятия интонаций русского языка в норме и при заболевании шизофренией отмечено, что у обследуемых пациентов имеется картина сглаженности функциональной асимметрии мозга, в то время как у здоровых испытуемых показано доминирование правого полушария.
Несмотря на сложившиеся представления о специализации полушарий, согласно которым высокая вербализация и концентрация внимания, абстрагирование сопровождаются активацией левой гемисферы, а реализация конкретно-образного мышления, эмоциональная деятельность — относительно большей активацией правой [33], функциональная специфичность полушарий не носит абсолютного характера. Значение имеют межполушарные когерентные связи, изменяющиеся определенным образом при мыслительной деятельности, а также претерпевающие перестройки при очаговых поражениях мозга разной локализации [6].
В работе М.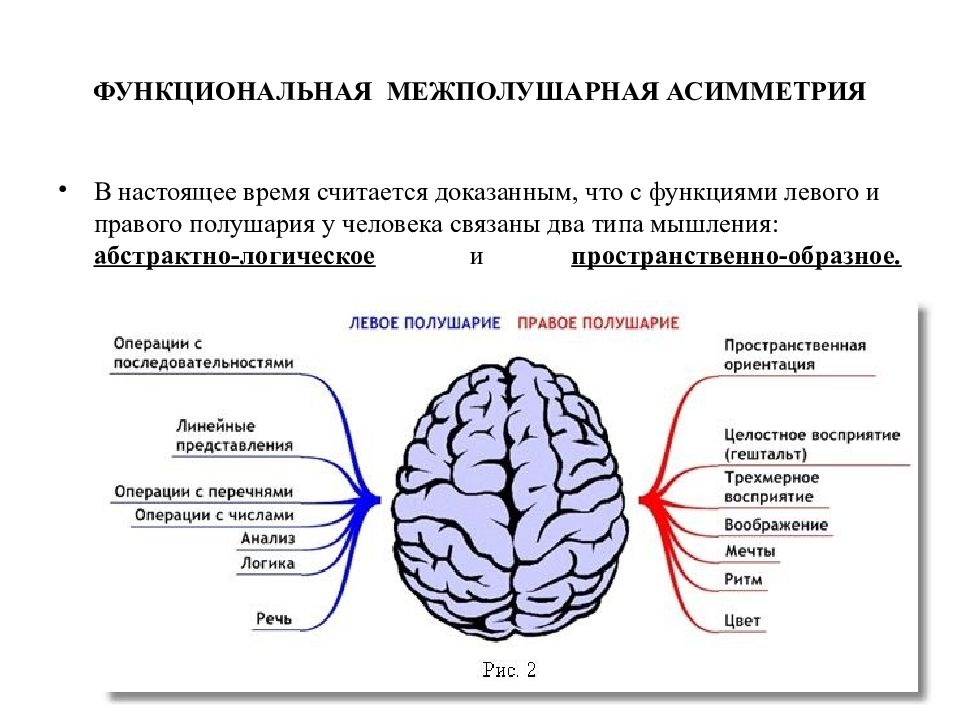 Н. Русаловой и соавт. [26] показано, что в процесс распознавания эмоций в речи вовлечены оба полушария. Результаты, полученные в клинике локальных поражений мозга, также позволяют сделать вывод, что за понимание эмоционального выражения речи отвечают не только височные области правой гемисферы, но и речевые центры в левой [39]. Значительное же увеличение в передних отделах левого полушария когерентности в гамма-диапазоне, отражающем когнитивные процессы, свидетельствует о более активном включении вербально-логических процессов при опознании и может явиться фактором, затрудняющим чувственно-непосредственное восприятие эмоций у лиц, плохо распознающих эмоциональные оттенки речи [26].
Н. Русаловой и соавт. [26] показано, что в процесс распознавания эмоций в речи вовлечены оба полушария. Результаты, полученные в клинике локальных поражений мозга, также позволяют сделать вывод, что за понимание эмоционального выражения речи отвечают не только височные области правой гемисферы, но и речевые центры в левой [39]. Значительное же увеличение в передних отделах левого полушария когерентности в гамма-диапазоне, отражающем когнитивные процессы, свидетельствует о более активном включении вербально-логических процессов при опознании и может явиться фактором, затрудняющим чувственно-непосредственное восприятие эмоций у лиц, плохо распознающих эмоциональные оттенки речи [26].
Результаты другого исследования [34] показали, что нейрофизиологические процессы, лежащие в основе обеспечения вербально-мыслительной деятельности, направленной на синтез отдельных слов и предложений, также протекают при высоком уровне системной взаимосвязи биоэлектрической активности коры обоих полушарий, особенно их височных и нижнелобных отделов. При этом в процесс реализации тесных межполушарных взаимодействий вовлечены
При этом в процесс реализации тесных межполушарных взаимодействий вовлечены
не только зона Брока и Вернике, но и симметричные им отделы правой гемисферы.
Интеграция правого и левого полушарий мозга необходима и для успешного творчества. Так, по данным А. Ю. Каразаевой и О. М. Разумниковой [17], повышение вербальной оригинальности сопровождается ослаблением полушарной асимметрии. Это обусловлено тем, что доминирование функций правого полушария является первостепенным условием создания оригинального образа, тогда как для построения оригинального по смыслу предложения требуется объединение возможностей обоих полушарий: уникальных для формирования отдаленных вербальных ассоциаций правого и использования определенных правил составления предложения, присущих левому.
Анатомическим субстратом межполушарного взаимодействия являются многочисленные мозговые комиссуры, однако ведущая роль принадлежит мозолистому телу. Одной из важных функций данной морфологической структуры является обеспечение возможности межполушарного торможения для дифференциации активности полушарий и более эффективной обработки информации. При повышении трудности выполняемого задания возрастает роль межполушарного взаимодействия. От качества межполушарного взаимодействия зависит и уровень интеллекта [18]. Межполушарная дезинтеграция, проявляющаяся в снижении межполушарной когерентности электроэнцефалограммы (ЭЭГ), коррелирует с уменьшением размеров мозолистого тела и приводит к ухудшению показателей когнитивной функции [44]. Как показали результаты магнитно-резонансной томографии, посттравматические стрессовые расстройства, сопровождающиеся высоким уровнем кортизола, также приводят к уменьшению размеров мозолистого тела и снижению межполушарного взаимодействия [50].
Одной из важных функций данной морфологической структуры является обеспечение возможности межполушарного торможения для дифференциации активности полушарий и более эффективной обработки информации. При повышении трудности выполняемого задания возрастает роль межполушарного взаимодействия. От качества межполушарного взаимодействия зависит и уровень интеллекта [18]. Межполушарная дезинтеграция, проявляющаяся в снижении межполушарной когерентности электроэнцефалограммы (ЭЭГ), коррелирует с уменьшением размеров мозолистого тела и приводит к ухудшению показателей когнитивной функции [44]. Как показали результаты магнитно-резонансной томографии, посттравматические стрессовые расстройства, сопровождающиеся высоким уровнем кортизола, также приводят к уменьшению размеров мозолистого тела и снижению межполушарного взаимодействия [50].
Результаты комплексного клинико-нейропсихологи-ческого обследования до и после операции пациентов с артериовенозными мальформациями мозолистого тела [7] не показали признаков левополушарной дисфункции при повреждении мозолистого тела. А вот функции правого полушария в сфере эмоций, восприятия, пространственной деятельности были нарушены. Правая гемисфера имеет очень тесные функциональные связи с левой, даже выполнение функций, в которых ведущую роль играет эта гемисфера, может нарушаться при повреждении мозолистого тела. Правое полушарие объединяет импульсы с обеих сторон пространства. Оно может оодновременно обрабатывать много разнообразной информации, т. е. способно рассматривать проблему в целом без разложения ее на составляющие и анализ. Однако для формирования представления как об отдельных объектах, так и об определенных видах деятельности необходим тесный контакт с «осознающим» левым полушарием [7].
А вот функции правого полушария в сфере эмоций, восприятия, пространственной деятельности были нарушены. Правая гемисфера имеет очень тесные функциональные связи с левой, даже выполнение функций, в которых ведущую роль играет эта гемисфера, может нарушаться при повреждении мозолистого тела. Правое полушарие объединяет импульсы с обеих сторон пространства. Оно может оодновременно обрабатывать много разнообразной информации, т. е. способно рассматривать проблему в целом без разложения ее на составляющие и анализ. Однако для формирования представления как об отдельных объектах, так и об определенных видах деятельности необходим тесный контакт с «осознающим» левым полушарием [7].
Нейрофизиологическая организация функциональной межполушарной асимметрии головного мозга
определяется конституциональными и генетическими различиями. Так, среди коренных народов Севера преобладают левши и амбидекстры в сравнении с русскими, проживающими там же. Проявляющийся при этом характер межполушарных отношений обусловлен не только климатическими условиями проживания, но и особенностями культурной среды соответствующей этнической группы. Например, среди японских школьников с учетом всех переученных около 11 % предпочитают использовать левую руку [2].
Проявляющийся при этом характер межполушарных отношений обусловлен не только климатическими условиями проживания, но и особенностями культурной среды соответствующей этнической группы. Например, среди японских школьников с учетом всех переученных около 11 % предпочитают использовать левую руку [2].
Существуют известные представления о том, что право- и леворукие различаются по объему мозга в полушариях: у праворуких лиц левое полушарие имеет тенденцию быть больше, особенно в зонах Брока и Вернике. У левшей предполагается частичная утрата генетического контроля над образованием функциональной асимметрии, мозг таких людей более симметричен [31].
Л. А. Жаворонкова [14], проанализировав ЭЭГ здоровых лиц в разных состояниях (спокойное бодрствование, активная деятельности и засыпание), выявила особенности межполушарной асимметрии правшей и левшей как отражение взаимодействия коры и регуляторных систем мозга. Полученные данные свидетельствуют о большей функциональной связи левого полушария с активирующими системами ствола у правшей. В отличие от этого у левшей подкорковые регуляторные системы оказывают на кору больших полушарий более диффузное влияние, с меньшим проявлением асимметрии. При этом, однако, у правшей отмечается конкурентное взаимодействие корковых и диэнцефальных структур, отражающееся в реципрок-ном характере изменения когерентности альфа- и тета-диапазонов в зоне корковой проекции диэнцефальных структур. У левшей же преобладает подкрепляющее взаимодействие, что находит отражение в синфазном изменении описанных выше ритмических составляющих ЭЭГ. Эти особенности организации мозга левшей могут быть основой успешного и плодотворного осуществления ими творческих видов деятельности, что подтверждается высокой численностью левшей среди музыкантов, художников, архитекторов [49].
Полученные данные свидетельствуют о большей функциональной связи левого полушария с активирующими системами ствола у правшей. В отличие от этого у левшей подкорковые регуляторные системы оказывают на кору больших полушарий более диффузное влияние, с меньшим проявлением асимметрии. При этом, однако, у правшей отмечается конкурентное взаимодействие корковых и диэнцефальных структур, отражающееся в реципрок-ном характере изменения когерентности альфа- и тета-диапазонов в зоне корковой проекции диэнцефальных структур. У левшей же преобладает подкрепляющее взаимодействие, что находит отражение в синфазном изменении описанных выше ритмических составляющих ЭЭГ. Эти особенности организации мозга левшей могут быть основой успешного и плодотворного осуществления ими творческих видов деятельности, что подтверждается высокой численностью левшей среди музыкантов, художников, архитекторов [49].
По мнению Л. А. Жаворонковой [14], особенности асимметрии мозга левшей не являются зеркальным отражением организации мозга правшей, а представляют собой альтернативный принцип его функционирования.
Особенности латерального устройства мозга во многом предопределяют и способность индивидуума к получению высоких результатов в определенном виде спорта. Периферическое отражение функциональной межполушарной специфики в виде индивидуального профиля асимметрии является фактором естественного отбора личностей, успешнее повышающих свою спортивную квалификацию, легче переносящих перенапряжение во время соревнований, быстрее и надежнее адаптирующихся к высоким психофизическим нагрузкам в жестко регламентируемых или, наоборот, ситуативных условиях [5, 16].
Так, Е. М. Бердичевская и соавт. [5], изучая индивидуальный профиль асимметрии (по схеме рука — нога — глаз) у стрелков и гандболистов как представителей видов спорта с принципиально различной структурой двигательной активности, определили наличие правосторонних предпочтений у спортсменов. Для стрелков характерно доминирование правой руки, ноги и глаза, которое авторы объясняют спецификой позы изготовки стрелка и конструкцией оружия.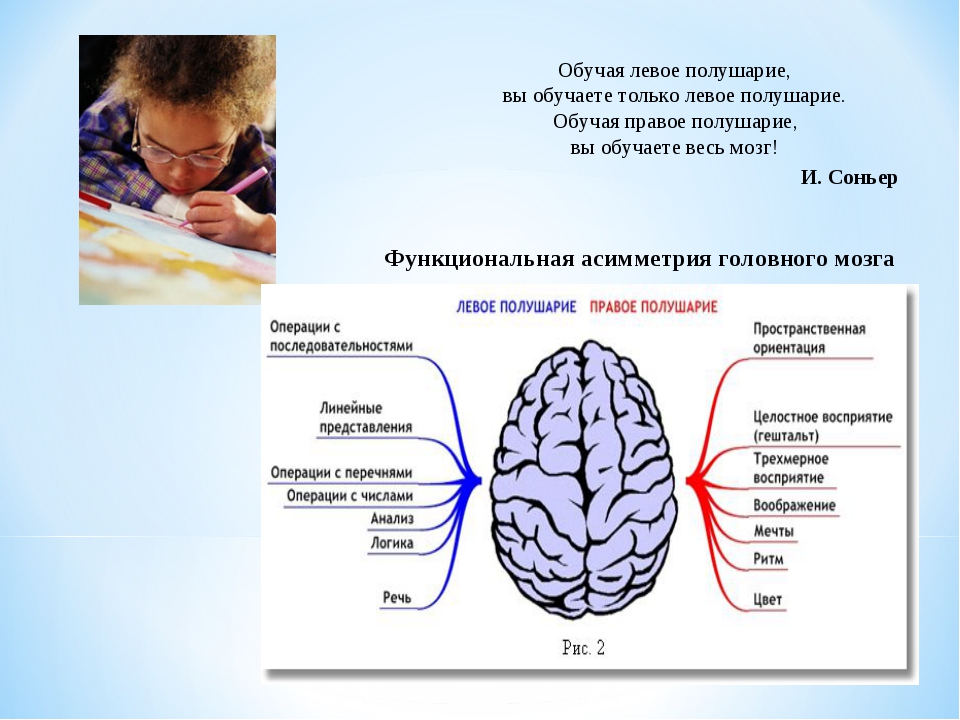 Гандболистам же свойственно парциальное распределение функций, что объясняется необходимостью билатерального восприятия пространства во время игры. Вовлечение в процесс правого полушария обеспечивает не только оценку текущей ситуации, но предвидение возможных ее изменений.
Гандболистам же свойственно парциальное распределение функций, что объясняется необходимостью билатерального восприятия пространства во время игры. Вовлечение в процесс правого полушария обеспечивает не только оценку текущей ситуации, но предвидение возможных ее изменений.
При изучении индивидуального профиля асимметрии у высококвалифицированных баскетболистов [30] особое внимание привлекает высокий процент леворуких спортсменов. Они наиболее часто встречаются среди занимающихся ситуационными видами спорта, для которых характерен прямой контакт между соперниками, а также в видах спорта, связанных с конкурентоспособной деятельностью рук.
Спортивная востребованность левшей может быть связана не только с особенностями их двигательного развития, но и со спецификой восприятия информации, стратегии мышления и стиля действия. Так, среди финалистов крупнейших международных соревнований по фехтованию число леворуких в 10 раз больше, чем в популяции. При этом левши в отличие от правшей, использующих контратакующий стиль, выполняют простые и быстрые действия, тем самым предпочитая атакующую манеру ведения боя [21].
При этом левши в отличие от правшей, использующих контратакующий стиль, выполняют простые и быстрые действия, тем самым предпочитая атакующую манеру ведения боя [21].
Таким образом, по мнению Ю. Н. Кабанова [16], определение профиля латеральной организации мозга служит признаком результативности во многих видах спорта и должно использоваться в процессе тренировок для более эффективного и гармоничного развития физических способностей спортсменов.
Сравнительный анализ показателей социально-психологической адаптации и межполушарной специализации мозга при помощи коэффициента ранговой корреляции Спирмена позволил выделить ряд значимых связей [4]. Авторы выяснили, что лица с доминированием левого полушария в стрессовых условиях принимают решение о возможности позитивного изменения ситуации и мобилизуют все свои ресурсы для его достижения. Индивидуумы с право-латеральной специализацией при столкновении со стрессовой ситуацией стремятся к уходу от проблемы физически или психически [20].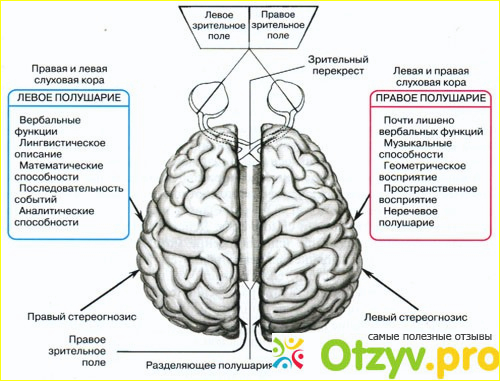 Полученные данные авторы объясняют индивидуальной гетерогенностью распределения нейромедиаторных систем головного мозга человека, которые в конечном итоге формируют психофизиолого-соматовегетативный статус личности, а также определяют характер межполушарной асимметрии. Лица с доминированием правого полушария характеризуются более высокой тревожностью эмоционально-вегетативного типа. Доминирование
Полученные данные авторы объясняют индивидуальной гетерогенностью распределения нейромедиаторных систем головного мозга человека, которые в конечном итоге формируют психофизиолого-соматовегетативный статус личности, а также определяют характер межполушарной асимметрии. Лица с доминированием правого полушария характеризуются более высокой тревожностью эмоционально-вегетативного типа. Доминирование
левого полушария показывает обратные процессы, т. е. высокую самооценку здоровья. По данным ряда авторов, у людей с доминирование левого полушария низкая склонность к депрессии, в отличии от право-полушарных [47].
И. И. Черемушникова [35], рассматривая связь полушарной асимметрии мозга и индивидуальных агрессивных стилей эмоционального поведения у первокурсников в период первых месяцев адаптации обучения, выявила, что у студентов с доминированием левополушарной активности (правый моторный профиль) преобладал положительный эмоциональный фон, а у людей с доминированием правополушарной активности — негативные эмоциональные состояния и отмечалась склонность к агрессии.
А. А. Земляной [15] были получены данные о различном влиянии прогредиентности заболевания эпилепсия на тип психической патологии у лиц с правым и левым профилями сенсомоторной асимметрии. У пациентов с правым профилем сенсомоторной асимметрии преимущественно страдали когнитивные и исполнительные функции, тогда как у пациентов с левым профилем сенсомоторной асимметрии в большей мере появлялись психопатологические феномены. E. Cubo et al. [43] также была отмечена тенденция к более глубокому ухудшению познавательной деятельности у пациентов с правосторонним нарушением моторной функции (патология левого полушария), особенно для внимания, исполнительной функции и памяти обучения.
Другой вопрос, имеющий отношение к генетике, связан с неодинаковой латерализацией мужского и женского мозга. С нейрофизиологических позиций появляется все больше данных, свидетельствующих о половых различиях в его структурной и нейрохимической организации. Мозг у мужчин больше, чем у женщин, особенно в лобной и затылочной зонах коры, тогда как у женщин сравнительно больше толщина коры в задней височной части нижней теменной области. Имеются данные, указывающие на разницу в толщине волокон передней части мозолистого тела, которые имеют более диффузную организацию проекций в правом полушарии у мужчин [48], тогда как задняя часть мозолистого тела оказывается больше у женщин. Существуют доказательства о связи уровня интеллекта у мужчин в большей степени с объемом серого вещества, а у женщин — белого [45]. Когнитивная деятельность разных видов, в том числе решение творческих задач, как образной, так и вербальной природы сопровождается разными формами взаимодействия полушарий у мужчин и женщин [41].
Мозг у мужчин больше, чем у женщин, особенно в лобной и затылочной зонах коры, тогда как у женщин сравнительно больше толщина коры в задней височной части нижней теменной области. Имеются данные, указывающие на разницу в толщине волокон передней части мозолистого тела, которые имеют более диффузную организацию проекций в правом полушарии у мужчин [48], тогда как задняя часть мозолистого тела оказывается больше у женщин. Существуют доказательства о связи уровня интеллекта у мужчин в большей степени с объемом серого вещества, а у женщин — белого [45]. Когнитивная деятельность разных видов, в том числе решение творческих задач, как образной, так и вербальной природы сопровождается разными формами взаимодействия полушарий у мужчин и женщин [41].
ЭЭГ-исследование праворуких мужчин и женщин [9] показало, что при воспроизведении вербальной информации когерентность биопотенциалов тета2-диапазона, связанного с процессами внимания, была выше у мужчин в правом полушарии. В левом полушарии когерентность альфа 1,2-ритмов, связанных с дифференцированной обработкой информации различной сенсорной модальности, больше возрастала у
В левом полушарии когерентность альфа 1,2-ритмов, связанных с дифференцированной обработкой информации различной сенсорной модальности, больше возрастала у
женщин. Только у женщин обнаружены латеральные различия в реактивности внутриполушарной когерентности для вышеуказанных диапазонов, обусловленные снижением когерентных взаимодействий в правом и повышением — в левом полушарии. Выявленные половые различия мощности и когерентности ЭЭГ связаны с использованием разных стратегий запоминания информации: преимущественно эпизодической памяти — мужчинами и семантического полимодального кодирования — женщинами.
В исследованиях И. В. Тарасовой и соавт. [29] по изучению частотно-пространственной организации активности коры при выполнении образной творческой задачи лицами разных полов наблюдалась большая десинхронизации тета1-ритма в правом в сравнении с левым полушарием у женщин, у мужчин асимметрия не обнаруживалась.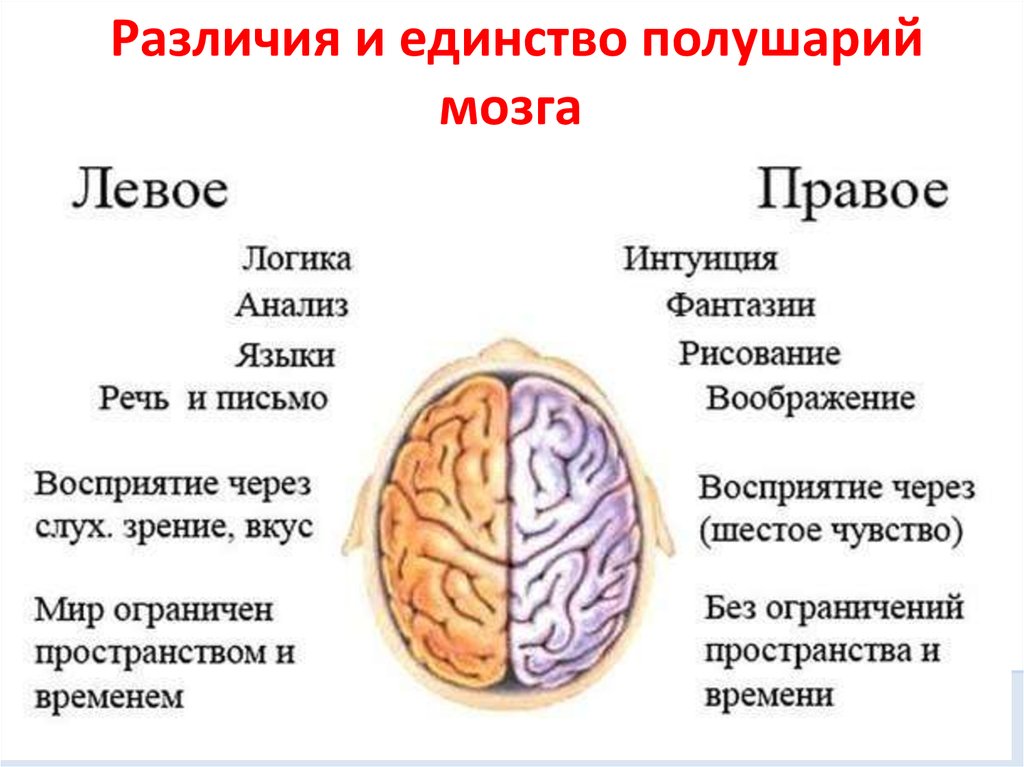 Достоверно менее выраженная десинхронизация тета1-ритма в левом полушарии свидетельствует о привлечении женщинами левополушарных стратегий при образном креативном мышлении.
Достоверно менее выраженная десинхронизация тета1-ритма в левом полушарии свидетельствует о привлечении женщинами левополушарных стратегий при образном креативном мышлении.
Согласно представлениям О. М. Разумниковой и соавт. [23] связанные с выполнением заданий изменения альфа2-ритма в большей мере характерны при вербальном творческом мышлении женщин, а бета2-ритма при образном — мужчин. Эти данные служат еще одним доказательством существования половых различий в самих механизмах деятельности мозга при отсутствии таких различий на поведенческом уровне, показанном при исследовании когнитивных функций у мужчин и женщин [40].
Гендерные различия электрической активности мозга обнаружены не только у здоровых мужчин и женщин, но и при патологии. Так, при хроническом алкоголизме изменения спектрально-когерентных характеристик ЭЭГ характеризуются у женщин большим, чем у мужчин, повышением синхронизации низких частот (дельта, тета) и снижением высоких — бета 1 и бета 2.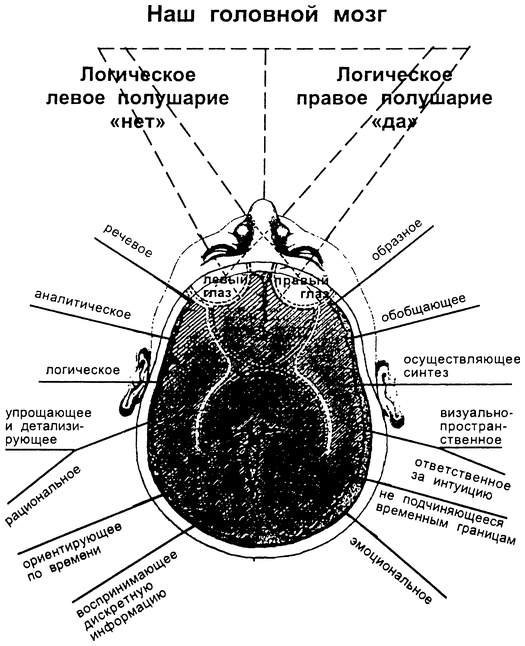 Подобные изменения отражают специфику изменений функционального состояния мозга у мужчин и женщин при алкоголизме [10].
Подобные изменения отражают специфику изменений функционального состояния мозга у мужчин и женщин при алкоголизме [10].
Все вышеперечисленное согласуется с классической концепцией о стабильной доминантности одного полушария или какого-либо участка полушария. Однако согласно современным представлениям одним из свойств функциональной межполушарной асимметрии является переключаемость, которая срабатывает при изменении функционального состояния организма для адекватного соответствия деятельности головного мозга новым условиям. Некоторые исследователи отмечают, что динамика межполушарной асимметрии меняется лишь при значимых сдвигах функциональных состояний, например таких, как переход от спокойного состояния к стрессу, при этом полушарное доминирование может снижаться вплоть до инверсии [31]. Изменение уровня бодрствования является естественным процессом последовательного изменения функционального состояния. В работе М. Н. Русаловой [25] анализ показателей ЭЭГ правшей показал, что в активном
Н. Русаловой [25] анализ показателей ЭЭГ правшей показал, что в активном
состоянии сознания межполушарные связи в высокочастотных диапазонах — бета-2 и гамма — легче обнаруживаются в левом полушарии мозга. В то же время для состояния торможения «внутренней речи», т. е. в содержании сознания отсутствует оформленная в словах мыслительная активность (самый низкий уровень бодрствования), характерно перемещение максимума большинства когерентных связей в правое полушарие, за исключением дельта-диапазона. Меж-полушарная динамика амплитудных автоспектров и фокусов когерентных связей подтверждает представление о том, что характер межполушарной асимметрии биоэлектрической активности мозга зависит от его функционального состояния.
Анализ результатов исследования особенностей структуры поля биопотенциалов головного мозга в различные периоды естественного ночного сна [38] выявил существенные различия в системной деятельности левого и правого полушарий. Полученные данные указывают на значимое возрастание в начале периода сна уровня внутриполушарных связей ЭЭГ как в левом, так и особенно в правом полушарии. При дальнейшем углублении сна и переходе к мед-ленноволновым стадиям доминирование правого полушария меняется на более выраженное и стойкое усиление внутриполушарных связей ЭЭГ в пределах левого полушария. Преимущественная доминантность левого полушария имеет тенденцию к сохранению и в период парадоксального сна.
Полученные данные указывают на значимое возрастание в начале периода сна уровня внутриполушарных связей ЭЭГ как в левом, так и особенно в правом полушарии. При дальнейшем углублении сна и переходе к мед-ленноволновым стадиям доминирование правого полушария меняется на более выраженное и стойкое усиление внутриполушарных связей ЭЭГ в пределах левого полушария. Преимущественная доминантность левого полушария имеет тенденцию к сохранению и в период парадоксального сна.
Динамичность межполушарной асимметрии обнаруживается также и в условиях патологии. Так, в работе С. Г. Александрова [3] показано, что у лиц с опийной зависимостью подавление активности левой гемисферы, связанной с дофаминергической системой, приводит к активации правого полушария, ответственного за формирование новых адаптивных программ. В условиях же отмены наркотического вещества и купирования абстинентного синдрома начинается нормализация нарушенного баланса межполушарной активации. Растормаживание левого полушария в результате нормализации функционального состояния дофаминергической системы приводит к сопряженному снижению активности правого полушария. Последнее выражается в более благоприятной динамике нормализации психофизиологических функций у пациентов с доминированием левого полушария.
Растормаживание левого полушария в результате нормализации функционального состояния дофаминергической системы приводит к сопряженному снижению активности правого полушария. Последнее выражается в более благоприятной динамике нормализации психофизиологических функций у пациентов с доминированием левого полушария.
Исследование особенностей межполушарной асимметрии у лиц мужского пола [19] позволило выявить, что в состоянии покоя у мужчин с высоким уровнем деструктивности фокус межполушарной асимметрии расположен в височных и фронтальных отделах левого, у лиц с низким уровнем деструктивности — в аналогичных отделах правого полушария. Моделирование агрессивной деятельности в обеих группах вызывало у обследуемых смещение фокуса активности в правое полушарие, независящее от исходной направленности асимметрии.
К факторам, определяющим динамику межполу-шарной асимметрии, также относятся и изменения
окружающей среды, влияющие на функциональное состояние организма. Так, анализ особенностей межполушарных взаимодействий мозга в процессе регуляции организационно-временной структуры организма в острой и скрытой фазе десинхроноза, вызванного авиаперелетом на 7 часовых поясов [12], показал, что в фазе острой адаптации (нарушение ритма сон — бодрствование) основную нагрузку по регуляции функций несет правое полушарие. В этот период испытуемые демонстрируют наименьшее количество ошибок (по сравнению с состоянием до перелета), чему сопутствует большая асимметрия. Данный факт авторы объясняют включением дополнительных механизмов адаптации, которые выводят организм из кризисного состояния. В фазе скрытого десинхроноза (в «доводке» инертных функций) преобладает регулирующая роль левополушарной активности. Правое полушарие мозга в основном обеспечивает формирование новой программы взаимодействия с изменившимися условиями окружающей среды. В свою очередь, улучшение воспроизведения новой информации происходит преимущественно в левом полушарии. С восстановлением функциональных показателей и циркадианной организации биоритмов обнаруживается восстановление исходных межполу-шарных взаимоотношений.
Так, анализ особенностей межполушарных взаимодействий мозга в процессе регуляции организационно-временной структуры организма в острой и скрытой фазе десинхроноза, вызванного авиаперелетом на 7 часовых поясов [12], показал, что в фазе острой адаптации (нарушение ритма сон — бодрствование) основную нагрузку по регуляции функций несет правое полушарие. В этот период испытуемые демонстрируют наименьшее количество ошибок (по сравнению с состоянием до перелета), чему сопутствует большая асимметрия. Данный факт авторы объясняют включением дополнительных механизмов адаптации, которые выводят организм из кризисного состояния. В фазе скрытого десинхроноза (в «доводке» инертных функций) преобладает регулирующая роль левополушарной активности. Правое полушарие мозга в основном обеспечивает формирование новой программы взаимодействия с изменившимися условиями окружающей среды. В свою очередь, улучшение воспроизведения новой информации происходит преимущественно в левом полушарии. С восстановлением функциональных показателей и циркадианной организации биоритмов обнаруживается восстановление исходных межполу-шарных взаимоотношений. Таким образом, увеличение функциональной асимметрии мозга коррелирует с повышением работоспособности человека.
Таким образом, увеличение функциональной асимметрии мозга коррелирует с повышением работоспособности человека.
Это подтверждают и результаты, полученные в исследованиях с участием праворуких лиц в процессе работы на компьютере [11]. У людей, не имеющих навыка операторской работы на компьютере, ведущую роль в мозговом обеспечении исследуемой деятельности играет левое полушарие. Отсутствие навыка требует определенных усилий, что, несомненно, и отражается на межполушарном взаимодействии, когда каждое полушарие, функционируя согласно своей специализации, вносит свой вклад в процесс реализации деятельности, выполняемой впервые. С упрочнением навыка выполнение задания приобретает стереотипный характер, что приводит к нивелированию межполушарной разницы у «профессионалов».
Таким образом, электрофизиологические и нейрохимические признаки функциональной межполу-шарной асимметрии, трудно идентифицируемые в состоянии покоя, контрастируются формирующимися функциональными системами. В этом плане функциональные системы женской репродукции обеспечивают естественную, долговременную и латерализован-ную «нагрузку» на мозг. Данные, полученные при обследовании беременных в сроке 35—36 недель [8], свидетельствуют о том, что для неосложненной беременности характерно превалирование активации ЭЭГ в центральных и височных областях полушария, контралатерального стороне расположения плаценты в полости матки. Ипсилатеральная исходно-доминантному полушарию, а значит, чаще левосторонняя плацентация, приводя к пространственному несоот-
В этом плане функциональные системы женской репродукции обеспечивают естественную, долговременную и латерализован-ную «нагрузку» на мозг. Данные, полученные при обследовании беременных в сроке 35—36 недель [8], свидетельствуют о том, что для неосложненной беременности характерно превалирование активации ЭЭГ в центральных и височных областях полушария, контралатерального стороне расположения плаценты в полости матки. Ипсилатеральная исходно-доминантному полушарию, а значит, чаще левосторонняя плацентация, приводя к пространственному несоот-
ветствию центральной и периферической асимметрии, характеризуется отсутствием признаков гестационной доминантны и становится одним из важнейших факторов угрозы прерывания беременности. Именно в таких случаях и наблюдается миграция плаценты, что способствует самопроизвольному ослаблению или исчезновению угрозы невынашивания.
В настоящее время предметом активного изучения является вопрос о возрастных изменениях межпо-лушарных взаимодействий, перестройках функциональной активности правого и левого полушарий головного мозга в процессе старения.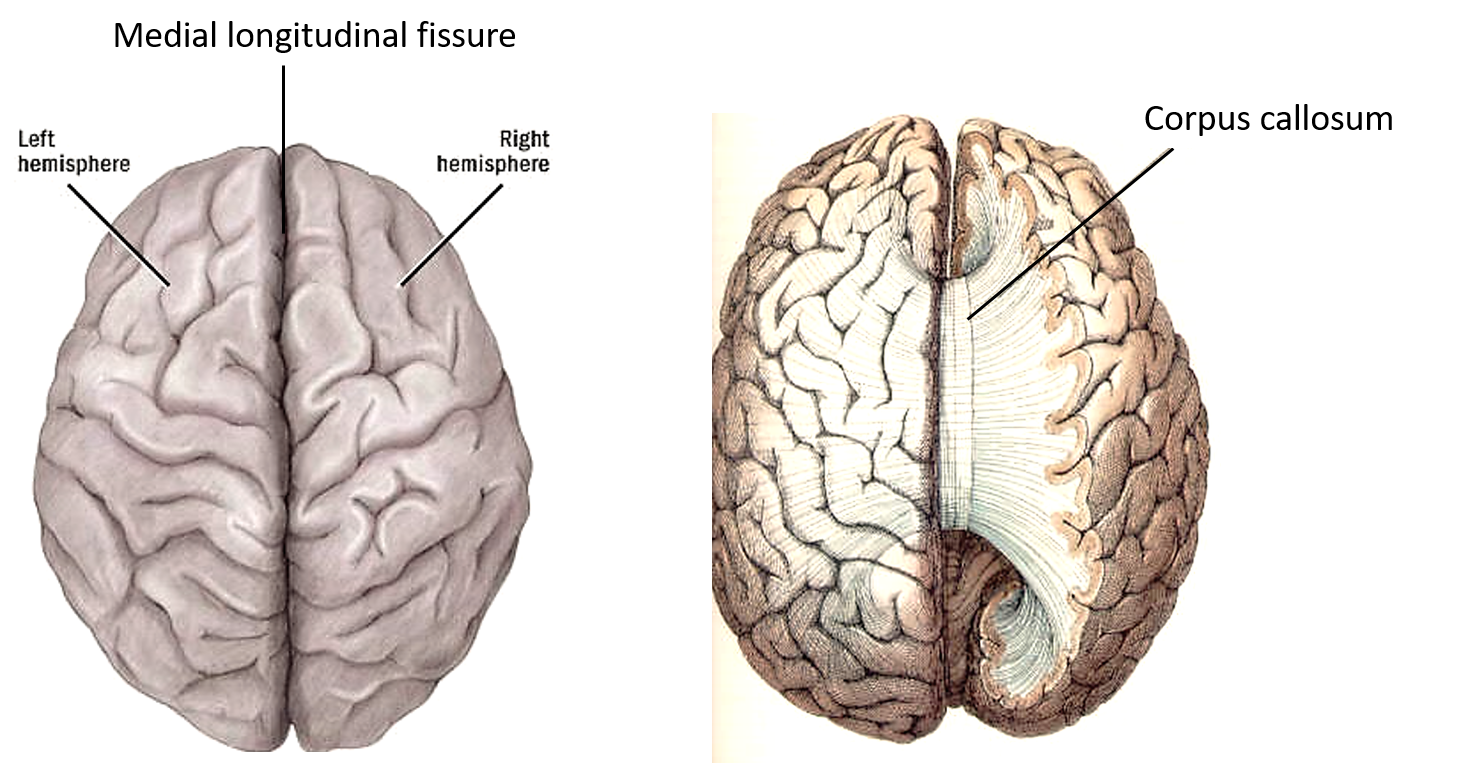 Существует мнение о том, что с возрастом происходит постепенное сглаживание межполушарных различий, что, возможно, является выражением пластичности и компенсаторным механизмом, предотвращающим функции мозга от дегенеративных изменений [46]. Так, результаты исследования Е. В. Севостьяновой [27] показали, что на Севере при наличии артериальной гипертензии имеет место не нормальное физиологическое старение, а патологическое, ускоренное, возникающее в значительно более молодом возрасте, чем в средних широтах, и, возможно, имеющее ряд специфических особенностей. У пришлых жителей Севера с артериальной гипертонией происходит достоверное снижение функциональной активности обоих полушарий головного мозга в возрасте 56— 65 лет и сглаживание функциональной межполушарной асимметрии после 45 лет. Е. В. Севостьянова [27] полагает, что в механизмах такого ускоренного старения человека на Севере существенную роль играют процессы снижения функциональной активности полушарий головного мозга и межполушарной дезинтеграции, связанные со стрессирующим экологическим воздействием.
Существует мнение о том, что с возрастом происходит постепенное сглаживание межполушарных различий, что, возможно, является выражением пластичности и компенсаторным механизмом, предотвращающим функции мозга от дегенеративных изменений [46]. Так, результаты исследования Е. В. Севостьяновой [27] показали, что на Севере при наличии артериальной гипертензии имеет место не нормальное физиологическое старение, а патологическое, ускоренное, возникающее в значительно более молодом возрасте, чем в средних широтах, и, возможно, имеющее ряд специфических особенностей. У пришлых жителей Севера с артериальной гипертонией происходит достоверное снижение функциональной активности обоих полушарий головного мозга в возрасте 56— 65 лет и сглаживание функциональной межполушарной асимметрии после 45 лет. Е. В. Севостьянова [27] полагает, что в механизмах такого ускоренного старения человека на Севере существенную роль играют процессы снижения функциональной активности полушарий головного мозга и межполушарной дезинтеграции, связанные со стрессирующим экологическим воздействием.
Многолетние клинические и психофизиологические исследования [13] участников ликвидации последствий аварии на Чернобыльской атомной электростанции (АЭС) выявили стойкие нарушения пространственно-временной организации ЭЭГ и когнитивных функций. Наряду с этим у ликвидаторов аварии было обнаружено уплощение ЭЭГ, нарастающее с течением времени. При этом выявлено двухфазное изменение процессов: на первых этапах развития заболевания (через 3—5 лет после воздействия радиации) преобладали процессы гиперсинхронизации (при участии диэнцефальных и лимбических структур мозга). В более отдаленные сроки после такого воздействия формировались процессы гипосинхронизации, особенно в лобных отделах, при возможном ослаблении взаимодействия с активирующими системами стволовых структур мозга. Подобная динамика является отражением включения компенсаторных процессов ЦНС в более ранние сроки после облучения малыми дозами радиации, с последующим их ослаблением и развитием процессов декомпенсации в более отдаленные сроки развития заболевания. Выявленные особенности изменения ЭЭГ и высших психических функций у участников ликвидации последствий аварии
Выявленные особенности изменения ЭЭГ и высших психических функций у участников ликвидации последствий аварии
на Чернобыльской АЭС свидетельствуют о раннем или преждевременном старении в результате воздействия радиации.
Таким образом, для конкретного функционального состояния существуют характерные закономерности формирования и организации межполушарных отношений, на основании чего были выдвинуты представления о динамических свойствах функциональной межполушарной асимметрии [31]. По мнению A. D. Craig [42], динамическая асимметрия в значительной мере связана с асимметрией вегетативной нервной системы. Основываясь на том, что стимуляция симметричных образований мозга, главным образом инсулярной коры, вызывает противоположные изменения вегетативных функций, он предположил, что симпатические и парасимпатические волокна, идущие из вентромедиального ядра тала-муса, перекрещиваются таким образом, что большая часть симпатических волокон направляется в правую инсулярную кору, а парасимпатических — в левую. Эта асимметрия вегетативной нервной системы предопределяет динамическую асимметрию как из-за разной активации симпатических и парасимпатических волокон в условиях покоя и стресса, так и из-за естественной суточной ритмики симпатического и парасимпатического тонуса, что подтверждается результатами исследований [22].
Эта асимметрия вегетативной нервной системы предопределяет динамическую асимметрию как из-за разной активации симпатических и парасимпатических волокон в условиях покоя и стресса, так и из-за естественной суточной ритмики симпатического и парасимпатического тонуса, что подтверждается результатами исследований [22].
Анализ выполнения когнитивной пробы беглости словесных ответов [32] позволил В.Ф. Фокину и соавт. предположить, что существует не только воздействие когнитивной деятельности на вегетативную регуляцию сердечно-сосудистой системы, но прямое или опосредованное влияние динамики вегетативного тонуса на некоторые мнестические и ассоциативные процессы в центральной нервной системе. Левое полушарие и парасимпатический отдел вегетативной нервной системы формируют структуры, образующие функциональную связь, посредством которой осуществляется регулирующее влияние нейрохимических процессов, лежащих в основе логического мышления, памяти и осознанных эмоций. Правое же полушарие и симпатический отдел формируют аналогичную связь, лежащую в основе процессов, регулирующих образное мышление, память и безотчетные эмоции [20].
Правое же полушарие и симпатический отдел формируют аналогичную связь, лежащую в основе процессов, регулирующих образное мышление, память и безотчетные эмоции [20].
В ряде исследований установлена зависимость параметров иммунной системы здоровых людей и пациентов с некоторыми иммунопатологическими заболеваниями [ 1 ] от особенностей функциональной межполушарной асимметрии головного мозга, влияние которой опосредуется через вегетативную нервную систему. Выявлены достоверные изменения параметров автономной нервной системы, свидетельствующие об истощении вегетативного обеспечения деятельности с относительным повышением симпатической активности на фоне снижения активности парасимпатического отдела. При этом определены разнонаправленные изменения параметров иммунной системы у пациентов с раком желудка с повышенной симпатической активностью в зависимости от характе-
ра функциональной межполушарной асимметрии [28].
Таким образом, афферентно-эфферентные связи мозга с различными системами организма предопределяют взаимовлияние центральных и периферических асимметрий, что находит непременный асимметричный отклик в сопряженных органах другого уровня [37].
Рассмотренные в данном обзоре результаты исследований, касающиеся стационарных и динамических свойств функциональной специализации полушарий, показали, что с позиций классических и современных представлений вопрос организации функциональной межполушарной асимметрии вызывает большой интерес у разных специалистов. Современный этап изучения функциональной асимметрии мозга представляет собой бурно развивающийся процесс, который приносит все новые открытия.
Список литературы
1. Абрамова Т. Я., Соловьева И. Г., Сизиков А. Э., Герцог О. А., Сулутьян А. Е., Коненкова Л. П., Шишкова И. В., Абрамов С. В., Кожевников В. С., Абрамов В. В. Психофизиологический иммунный статус больных ревматоидным артритом // Медицинская иммунология. 2008. Т. 10, № 1. С. 43-50.
2. Агаджанян Н. А., Макарова И. И. Этический аспект адаптационной физиологии и заболеваемости населения // Экология человека. 2014. № 3. С. 3-13.
3. Александров С. Г. Специфика межполушарных отношений у больных с опийной зависимостью в абстинентный и ранний постабстинентный период // Физиология человека. 2004. Т. 30, № 4. С. 40-45.
4. Антропова Л. К., Андроникова О. О., Куликов В. Ю., Козлова Л. А. Функциональная асимметрия мозга и индивидуальные психофизиологические особенности человека // Медицина и образование в Сибири. 2011. № 3. URL: http://www.ngmu.ru/cozo/mos/article/text_full.php?id=485 (дата обращения 06.06.2015).
5. Бердичевская Е. М., Гронская А. С., Хачатуро-ва И. Э. Специфика латерального фенотипа в стрельбе и гандболе // Физическая культура, спорт — наука и практика. 2009. № 3. С. 27-29.
6. Болдырева Г. Н., Шарова Е. В., Коптелов Ю. М., Щепетков А. Н., Никитин К. В., Корниенко В. Н., Фадеева Л. М. Исследование генеза патологических паттернов ЭЭГ при опухолевом и травматическом поражении мозга человека // Физиология человека. 2005. Т. 31, № 1. С. 24-32.
7. Буклина С. Б. Мозолистое тело, межполушарное взаимодействие и функции правого полушария мозга // Журнал неврологии и психиатрии. 2004. Т. 104, № 5. С. 8-14.
8. Васильева В. В. Пространственно-временная организация биоэлектрической активности мозга при гестационной доминанте // Журнал высшей нервной деятельности. 2007. Т. 57, № 3. С. 292-302.
9. Вольф Н. В., Разумникова О. М. Половые различия полушарных пространственно-временных паттернов ЭЭГ при воспроизведении вербальной информации // Физиология человека. 2004. Т. 30, № 3. С. 27-34.
10. Глазкова В. А., Свидерская Н. Е. Гендерные различия при хроническом алкоголизме по данным ЭЭГ // Журнал высшей нервной деятельности. 2006. Т. 56, № 5. С. 597-602.
11. Григорян В. Г., Агабян А. Р, Аракелян А. Н. Степа-нян А. Ю. Особенности межполушарных взаимоотношений при отсутствии и наличии навыка // Физиология человека. 2006. Т. 32, № 4. С. 134-137.
12. Ежов С. Н., Кривощеков С. Г. Особенности психомоторных реакций и межполушарных отношений мозга на этапах временной адаптации // Физиология человека. 2004. Т. 30, № 2. С. 53-57.
13. Жаворонкова Л. А., Белостоцкий А. П., Холодова Н. Б., Купцова С. В., Снегирева И. П., Куликов М. А., Окнина Л. Б. Нарушения высших психических функций и когнитивных слуховых вызванных потенциалов у ликвидаторов Чернобыльской аварии // Журнал неврологии и психиатрии им. С. С. Корсакова. 2012. № 5. С. 62-69.
14. Жаворонкова Л. А. Особенности межполушарной асимметрии ЭЭГ правшей и левшей как отражение взаимодействия коры и регуляторных систем мозга // Функциональная межполушарная асимметрия : хрестоматия. Под ред. Н. Н. Боголепова, В. Ф. Фокина. М. : Научный мир, 2004. С. 287-292.
15. Земляная А. А., Калинин В. В., Ковязина М. С., Крылов О. Е. Психические расстройства и сенсомоторная асимметрия головного мозга у больных эпилепсией // Журнал неврологии и психиатрии. 2010. Вып. 2. Т. 63, № 3. С. 70-76.
16. Кабанов Ю. Н. Успешность спортивной деятельности и функциональная асимметрия головного мозга // Мир науки, культуры, образования. 2009. № 3. С. 194-201.
17. Каразаева А. Ю., Разумникова О. М. Взаимосвязь креативности и полушарных процессов селекции информации: значение моторной асимметрии // Журнал высшей нервной деятельности. 2012. Т. 62, № 3. С. 279-285.
18. Ковязина М. С., Балашова Е. Ю. Межполушарное взаимодействие при нормальном и отклоняющемся развитии: мозговые механизмы и психологические особенности // Руководство по функциональной межполушарной асимметрии. М. : Научный мир, 2009. Гл. 7. С. 185-206.
19. Кожевников С. П., Проничев И. В. Изменение параметров межполушарной асимметрии при моделировании агрессивного поведения // Журнал высшей нервной деятельности. 2012. Т. 62, № 1. С. 5-11.
20. Куликов В. Ю., Антропова Л. К., Козлова Л. А. Влияние функциональной асимметрии мозга на стратегию поведения индивида в стрессовой ситуации // Медицина и образование в Сибири. 2010. № 5. URL: http://ngmu. ru/cozo/mos/article/text_full.php?id=452 (дата обращения 06.06.2015).
21. Москвина Н. В., Москвин В. А. Леворукость в спорте высших достижений // Спортивный психолог. 2010. Т. 20, № 2. С. 25-29.
22. Пестряев В. А., Сафина Т. В. Межполушарная асимметрия трофотропной и эрготропной регуляции // Асимметрия. 2014. Т. 8, № 2. С. 48-58.
23. Разумникова О. М., Вольф Н. В., Тарасова И. В. Стратегия и результат: половые различия в электрофизиологических коррелятах вербальной и образной креативности // Физиология человека. 2009. Т. 35, № 3. С. 31-41.
24. Ротенберг В. Межполушарная асимметрия, ее функция и онтогенез // Руководство по функциональной межполушарной асимметрии. М. : Научный мир, 2009. Гл. 6. С. 164-184.
25. Русалова М. Н. Асимметрия биоэлектрической активности мозга человека на разных уровнях сознания // Российский физиологический журнал им. И. М. Сеченова. 2006. Т. 92, № 12. С. 1393-1403.
26. Русалова М. Н, Кислова О. О., Образцова Л. В. Электрофизиологические показатели распознавания эмоций в речи // Успехи физиологических наук. 2010. Т. 42, № 2. С. 57-82.
27. Севостьянова Е. В. Особенности функциональной межполушарной асимметрии у жителей севера с артериальной гипертонией различного возраста // Материалы Всероссийской конференции «Современные направления исследований функциональной межполушарной асимметрии и пластичности мозга», Москва, 2-3 декабря, 2010. URL: http://www.cerebral-asymmetry.ru/3Conf/30 (дата обращения 03.06.2015).
28. Стенина А. С., Абрамова Т. Я., Соловьева И. Г., Кожевников В. С., Абрамов В. В. Вегетативный баланс и параметры иммунной системы у больных раком желудка с различным характером функциональной межполушарной асимметрии // Материалы Всероссийской конференции «Современные направления исследований функциональной межполушарной асимметрии и пластичности мозга», Москва, 2-3 декабря, 2010. URL: http://www.cerebral-asymmetry.ru/3Conf/30 (дата обращения 03.06.2015).
29. Тарасова И. В., Вольф Н. В., Разумникова О. М. Изменения мощности ЭЭГ при образном креативном мышлении у мужчин и женщин // Журнал высшей нервной деятельности. 2005. Т. 55, № 6. С. 762-767.
30. Тришин А. С., Тришин Е. С., Катрин Л. В., Бер-дичевская Е. М. Сравнительная характеристика профиля функциональной асимметрии у квалифицированных спортсменов в настольном теннисе и баскетболе // Физическая культура, спорт — наука и практика. 2012. № 4. С. 55-58.
31. Фокин В. Ф., Боравова А. И., Галкина Н. С., Пономарева Н. В., Шимко И. А. Стационарная и динамическая организация функциональной межполушарной асимметрии // Руководство по функциональной межполушарной асимметрии. М. : Научный мир, 2009. Гл. 14. С. 389-428.
32. Фокин В. Ф., Пономарева Н. В., Клопов В. И., Танашян М. М. Сосудистая реактивность, вызванная когнитивной нагрузкой, у больных дисциркуляторной энцефалопатией // Асимметрия. 2014. Т. 8, № 3. С. 4-22.
33. Фокин В. Ф. Эволюционный аспект центрально-периферической организации функциональной межполу-шарной асимметрии // Функциональная межполушарная асимметрия : хрестоматия / под ред. Н. Н. Боголепова, В. Ф. Фокина. М. : Научный мир, 2004. С. 47-79.
34. Цапарина Д. М., Цицерошин М. Н., Шеповальни-ков А. Н. Реорганизация межполушарного взаимодействия при речемыслительной деятельности, направленной на синтез слов и предложении // Физиология человека. 2007. Т. 33, № 1. С. 15-26.
35. Черемушникова И. И. Изучение влияния функциональной межполушарной асимметрии у студентов на склонность к агрессии, с последующей оценкой неспецифических адаптационных реакций организма // Асимметрия. 2011. Т. 5, № 3. С. 35-48.
36. Черниговская Т. В., Давтян С. Э, Петрова Н. Н., Стрельников К. Н. Специфика полушарной асимметрии восприятия интонаций в норме и при шизофрении // Физиология человека. 2004. Т. 30, № 4. С. 32-39.
37. Черноситов А. В., Орлов В. И., Васильева В. В. Функциональная межполушарная асимметрия мозга — как объект репродуктивного системогенеза // Руководство по функциональной межполушарной асимметрии. М. : Научный мир, 2009. Гл. 5. С. 145-163.
38. Шеповальников А. Н., Цицерошин М. Н., Зайцева Л. Г., Гальперина Е. И. Особенности системного
взаимодействия разных областей коры левого и правого полушарий мозга в различных стадиях сна у человека // Российский физиологический журнал им. И. М. Сеченова. 2012. Т. 98, № 10. С. 1228-1241.
39. Beaucousin V., Lacheret A., Turbelin M. R., Morel M., Mazoyer B., Tzourio-Mazoyer N. FMRI study of emotional speech comprehension // Cereb. Cortex. 2007. Vol. 17, N 2. Р. 339-352.
40. Bell E. C, Willson M. C, Wilman A. H, Dave S, Silverstone P. H. Males and females differ in brain activation during cognitive tasks // Neuroimage. 2006. Vol. 30, N 2. Р. 529-538.
41. Clements A. M, Rimrodt S. L, Abel J. R. Blankner J. G, Mostofsky S. H., Pekar J. J., Denckla M. B., CuttingL. E. Sex differences in cerebral laterality of language and visuospatial processing // Brain Lang. 2006. Vol. 98, N 2. Р. 150-158.
42. Craig A. D. Forebrain emotional asymmetry: a neuroanatomical basis? // Trends Cogn. Sci. 2005. Vol. 9, N 12. Р. 566-571.
43. Cubo Е. Martin P. M., Martin-Gonzalez J. A., Rodriguez-Blazquez C., Kulisevsky J. Motor laterality asymmetry and nonmotor symptoms in Parkinson’s // Movement disorders. 2010. Vol. 25, N 1. P. 70-75.
44. Francis A. N., Bhojraj T. S., Prasad K. M., Kulkarni S., Montrose D. M., Eack S. M., Keshavan M. S. Abnormalities of the corpus callosum in non-psychotic high-risk offspring of schizophrenia patients // Psychiatry Res. 2011. Vol. 191, N 1. Р. 9-15.
45. Haier R. J., Jung R. E, Yeo R. A. HeadK., Alkire M. T. The neuroanatomy of general intelligence: sex matters // Neuroimage. 2005. Vol. 25, N 1. Р. 320-327.
46. Hommet C., Destrieux C., Constans T., Berrut G. Aging and hemispheric cerebral lateralization // Psychol. Neuropsychiatr. Vieil. 2008. Vol. 6, N 1. Р. 49-56.
47. Logue D. D., Logue R. T., Kaufmann W. E. Belcher H. M. Psychiatric disorders and left-handedness in children living in an urban environment // Laterality. 2015. Vol. 20, N 2. Р. 249-256.
48. Luders E., Narr K. L., Zaidel E., Thompson P. M., Toga A. W. Gender effects on callosal thickness in scaled and unscaled space // Neuroreport. 2006. Vol. 17, N 11. Р. 1 103-1106.
49. Preti A., Vellante M. Creativity and psychopathology: higher rates of psychosis proneness and nonright-handedness among creative artists compared to same age and gender peers // J. Nerv. Ment. Dis. 2007. Vol. 195, N 10. Р. 837-845.
50. Villarreal G., Hamilton D .A., Graham D. P., Driscoll I., Qualls C., Petropoulos H., Brooks W. M. Reduced area of the corpus callosum in posttraumatic stress disorder // Psychiatry Res. 2004. Vol. 131, N 3. Р. 227-235.
References
1. Abramova T. Ya., Solov’eva I. G., Sizikov A. E., Gertsog O. A., Sulut’yan A. E., Konen-kova L. P., Shishkova I. V., Abramov S. V., Kozhevnikov V. S., Abramov V. V. Physiological immune status of patients with rheumatoid arthritis. Meditsinskaya immunologiya [Medical Immunology]. 2008, 10 (1), рр. 43-50. [in Russian]
2. Agadzhanjan N. A., Makarova I. I. Ethnic aspect of adaptive physiology and population morbidity. Ekologiya cheloveka [Human Ecology]. 2014, 3, рр. 3-13. [in Russian]
3. Aleksandrov S. G. Specificity of interhemispheric relations in patients with opioid dependence in withdrawal and early post-abstinent period. Fiziologiya cheloveka [Human Physiology]. 2004, 30 (4), рр. 40-45. [in Russian]
4. Antropova L. K., Andronikova O. O., Kulikov V. Yu., Kozlova L. A. Functional asymmetry of the brain and individual psycho-physiological characteristics of a man. Meditsina i obrazovanie v Sibiri [Health and education in Siberia]. 2011, 3. Available at: http://www.ngmu.ru/cozo/mos/article/ text_full.php?id=485 (accessed 06.06.2015).
5. Berdichevskaya E. M., Gronskaya A. S., Khachaturova I. E. The Specificity of the lateral phenotype in shooting and handball. Fizicheskaya kul’tura, sport — nauka i praktika [Physical Culture, Sport — Science and Practice]. 2009, 3, pp. 27-29. [in Russian]
6. Boldyreva G. N., Sharova E. V., Koptelov Yu. M., Shchepetkov A. N., Nikitin K. V., Kornienko V. N., Fadeeva L. M. The study of the Genesis of pathological EEG patterns in neoplastic and traumatic lesions of the human brain. Fiziologiya cheloveka [Human Physiology]. 2005, 31 (1), pp. 24-32. [in Russian]
7. Buklina S. B. Corpus callosum, interhemispheric interaction and function of the right hemisphere of the brain. Zhurnal nevrologii i psikhiatrii [Journal of Neurology and Psychiatry]. 2004, 104 (5), pp. 8-14. [in Russian]
8. Vasil’eva V. V. Spatial-temporal organization of brain activity during the gestational dominant Zhurn. vyssh. nervn. deyat. [Journal of Higher Nervous Activity]. 2007, 57(3), pp. 292-302. [in Russian]
9. Vol’f N. V., Razumnikova O. M. Sex differences hemispheric spatial-temporal EEG patterns during playback of verbal information. Fiziologiya cheloveka [Human Physiology]. 2004, 30 (3), pp. 27-34. [in Russian]
10. Glazkova V. A., Sviderskaya N. E. Gender differences in chronic alcoholism according to the EEG. Zhurn. vyssh. nervn. deyat. [Journal of Higher Nervous Activity]. 2006, 56 (5), pp. 597-602. [in Russian]
11. Grigoryan V. G., Agabyan A. R, Arakelyan A. N. Stepanyan A. Yu. Features of interhemispheric relations in the absence and presence of skill. Fiziologiya cheloveka [Human Physiology]. 2006, 32 (4), pp. 134-137. [in Russian]
12. Ezhov S. N., Krivoshchekov S. G. Features of psychomotor reactions and hemispheric relations of the brain at the stages of temporary adaptation. Fiziologiya cheloveka [Human Physiology]. 2004, 30 (2), pp. 53-57. [in Russian]
13. Zhavoronkova L. A., Belostotskii A. P., Kholodova N. B., Kuptsova S. V., Snegireva I. P., Kulikov M. A., Oknina L. B. Violations of higher mental functions and cognitive auditory evoked potentials in the liquidators of the Chernobyl accident. Zhurn. nevrologii i psikhiatrii im. S. S. Korsakova [Journal of Neurology and Psychiatry named S.S. Korsakoff’s]. 2012, 5, pp. 62-69. [in Russian]
14. Zhavoronkova L. A. Osobennosti mezhpolusharnoi asimmetrii EEG pravshei i levshei kak otrazhenie vzaimodeistviya kory i regulyatornykh sistem mozga [Features of interhemispheric asymmetry of the EEG righties and lefties as a reflection of the interaction of the cortex and regulatory systems of the brain]. Funktsional’naya mezhpolusharnaya asimmetriya: khrestomatiya [Functional hemispheric asymmetry: a reader]. Edited by N. N.Bogolepova, V. F. Fokin. Moscow, Scientific World. 2004, pp. 287-292.
15. Zemlyanaya A. A., Kalinin V. V., Kovyazina M. S., Krylov O. E. Mental disorders and sensorimotor asymmetry of the brain in patients with epilepsy. Zhurn. nevrologii i psikhologii [Journal of Neurology and Psychiatry]. 2010, iss. 2, 63 (3), pp. 70-76. [in Russian]
16. Kabanov Yu. N. The success of sports activities and functional asymmetry of the Mir nauki, kul’tury, obrazovaniya [The world of science, culture, education]. 2009, 3, pp. 194-
201. [in Russian]
17. Karazaeva A. Yu., Razumnikova O. M. The relationship of creativity and hemispheric processes of information selection: the value of motor asymmetry Zhurn. vyssh. nervn. deyat. [Journal of Higher Nervous Activity]. 2012, 62 (3), pp. 279285. [in Russian]
18. Kovyazina M. S., Balashova E. Yu. Interhemispheric interaction in normal and deviant development: brain mechanisms and psychological characteristics [Manual functional hemispheric asymmetry]. Moscow, Scientific World,
2009, pt.7, pp. 185-206.
19. Kozhevnikov S. P., Pronichev I. V. Changing the parameters of interhemispheric asymmetry in modeling of aggressive behavior. Zhurn. vyssh. nervn. deyat. [Journal of Higher Nervous Activity]. 2012, 62 (1), pp. 5-11. [in Russian]
20. Kulikov V. Yu., Antropova L. K., Kozlova L. A. Influence of functional asymmetry of the brain to the strategy of behavior of the individual in the stressful situation. Meditsina i obrazovanie v Sibiri [Health and education in Siberia]. 2010, 5. Available at: http://ngmu.ru/cozo/mos/article/text_full. php?id=452 (accessed 06.06.2015).
21. Moskvina N. V, Moskvin V. A. Left-handedness in high performance sport. Sportivnyi psikholog [Sport Psychologist].
2010, 20 (2), pp. 25-29. [in Russian]
22. Pestryaev V. A., Safina T. V. Hemispheric asymmetry trophotropic ergotropic and regulation. Asimmetriya [Asymmetry]. 2014, 8 (2), pp. 48-58. [in Russian]
23. Razumnikova O. M., Vol’f N. V., Tarasova I. V. Strategy and outcome: gender differences in electrophysiological correlates of verbal and imaginative creativity. Fiziologiya cheloveka [Human Physiology]. 2009, 35 (3), pp. 31-41. [in Russian]
24. Rotenberg V. Hemispheric asymmetry, its function and ontogeny [Mezhpolusharnaya asimmetriya, ee funktsiya i ontogenez]. Rukovodstvo po funktsional’noi mezhpolusharnoi asimmetrii [Manual functional hemispheric asymmetry]. Moscow, Scientific World, 2009, pt. 6, pp. 164-184.
25. Rusalova M. N. The asymmetry of the bioelectric activity of human brain at different levels of consciousness. Ros. fiziol. zhurnal im. I. M. Sechenova [Russian physiological journal named I. M. Setchenov]. 2006, 92 (12), pp. 13931403. [in Russian]
26. Rusalova M. N, Kislova O. O., Obraztsova L. V. Electrophysiological indices of recognition of emotions in speech. Uspekhi fiziologicheskih nauk [Advances in Physiological Sciences]. 2010, 42 (2), pp. 57-82. [in Russian]
27. Sevost’yanova E. V. Osobennosti funktsional’noi mezhpolusharnoi asimmetrii u zhitelei severa s arterial’noi gipertoniei razlichnogo vozrasta [Features of functional interhemispheric asymmetry of the residents of the North with arterial hypertension of different age]. Materialy Vserossiiskoi konferentsii «Sovremennye napravleniya issledovanii funk-tsional’noi mezhpolusharnoi asimmetrii i plastichnosti mozga», Moskva, 2-3 dekabrja 2010 [Proceedings of the conference «Modern lines of research of functional hemispheric asymmetry and brain plasticity», Moscow, 2-3 December 2010]. Moscow, 2010. Available at: http://www.cerebral-asymmetry.ru/3Conf/30 (accessed 03.06.2015).
28. Stenina A. S., Abramova T. Ya., Solov’eva I. G., Kozhevnikov V. S., Abramov V. V. Vegetativnyi balans i parametry immunnoi sistemy u bol’nykh rakom zheludka s razlichnym kharakterom funktsional’noi mezhpolusharnoi asimmetrii [Autonomic balance and parameters of the immune system in patients with gastric cancer with different nature of functional interhemispheric asymmetry]. Materialy
Vserossiiskoi konferentsii «Sovremennye napravleniya issledovanii funk-tsional’noi mezhpolusharnoi asimmetrii i plastichnosti mozga», Moskva, 2-3 dekabrja 2010 [Proceedings of the conference «Modern lines of research of functional hemispheric asymmetry and brain plasticity», Moscow, 2-3 December 2010]. Moscow, 2010. Available at: http://www.cerebral-asymmetry.ru/3Conf/30 (accessed 03.06.2015).
29. Tarasova I. V., Vol’f N. V., Razumnikova O. M. Changes in EEG power in a sort of creative thinking in men and women. Zhurn. vyssh. nervn. deyat. [Journal of Higher Nervous Activity]. 2005, 55 (6), pp. 762-767. [in Russian]
30. Trishin A. S., Trishin E. S., Katrich L. V., Berdichevskaya E. M. Comparative characteristics of the profile of functional asymmetry of the qualified athletes in table tennis and basketball. Fizicheskaya kul’tura, sport — nauka i praktika [Physical Culture, Sport — Science and Practice]. 2012, 4, pp. 55-58. [in Russian]
31. Fokin V. F., Boravova A. I., Galkina N. S., Ponomareva N. V., Shimko I. A. Statsionarnaya i dinamicheskaya organizatsiya funktsional’noi mezhpolusharnoi asimmetrii [Stationary and dynamic organization of functional interhemispheric asymmetry]. Rukovodstvo po funktsional’noi mezhpolusharnoi asimmetrii [Manual functional hemispheric asymmetry]. Moscow, Scientific World, 2009, ch. 14, pp. 389428.
32. Fokin V. F., Ponomareva N. V., Klopov V. I., Tanashyan M. M. Vascular reactivity caused by cognitive load, in patients with discirculatory encephalopathy. Asimmetriya [Asymmetry]. 2014, 8 (3), pp. 4-22. [in Russian]
33. Fokin V. F. Evolyutsionnyi aspekt tsentral’no-perifericheskoi organizatsii funktsional’noi mezhpolusharnoi asimmetrii [Evolutionary aspect of the Central-peripheral organization of the functional interhemispheric asymmetry]. Funktsional’naya mezhpolusharnaya asimmetriya: khrestomatiya [Functional hemispheric asymmetry: a reader]. Edited by N. N. Bogolepova, V. F. Fokin. Moscow, Scientific World. 2004, pp. 47-79.
34. Tsaparina D. M., Tsitseroshin M. N., Shepoval’nikov A. N. Russian A. N. Reorganization of interhemispheric interaction on rehabilitating activities aimed at the synthesis of words and sentence. Fiziologiya cheloveka [Human Physiology]. 2007, 33 (1), pp. 15-26. [in Russian]
35. Cheremushnikova I. I. Study of the effect of functional interhemispheric asymmetry of the students on the propensity to aggression, then assess nonspecific adaptive reactions of the organism. Asimmetriya [Asymmetry]. 2011, 5 (3), pp. 35-48. [in Russian]
36. Chernigovskaya T. V., Davtyan S. E, Petrova N. N., Strel’nikov K. N. Specifics of hemispheric asymmetry in the perception of intonation in normal and schizophrenia. Fiziologiya cheloveka [Human Physiology]. 2004, 30 (4), pp. 32-39. [in Russian]
37. Chernositov A. V., Orlov V. I., Vasil’eva V. V. Funktsional’naya mezhpolusharnaya asimmetriya mozga — kak ob»ekt reproduktivnogo sistemogeneza [Functional interhemispheric asymmetry of the brain as an object of the reproductive system Genesis]. Rukovodstvo po funktsional’noi mezhpolusharnoi asimmetrii [Manual functional hemispheric asymmetry]. Moscow, Scientific World, 2009, pt. 5, pp. 145-163.
38. Shepoval’nikov A. N., Tsitseroshin M. N., Zaitseva L. G.,
Gal’perina E. I. Peculiarities of the system of interaction between different areas of the cortex of the left and right hemispheres of the brain in various stages of sleep in humans. Ros. fiziol. zhurnal im. I. M. Sechenova [Russian physiological journal named I. M. Setchenov]. 2012, 98 (10), рр. 12281241. [in Russian]
39. Beaucousin V., Lacheret A., Turbelin M. R., Morel M., Mazoyer B., Tzourio-Mazoyer N. FMRI study of emotional speech comprehension. Cereb. Cortex. 2007, 17 (2), рр. 339352.
40. Bell E. C., Willson M. C., Wilman A. H., Dave S., Silverstone P. H. Males and females differ in brain activation during cognitive tasks. Neuroimage. 2006, 30 (2), рр. 529538.
41. Clements A. M., Rimrodt S. L., Abel J. R. Blank-ner J. G., Mostofsky S. H., Pekar J. J., Denckla M. B., Cutting L. E. Sex differences in cerebral laterality of language and visuospatial processing. Brain Lang. 2006, 98 (2), рр. 150-158.
42. Craig A. D. Forebrain emotional asymmetry: a neuroanatomical basis? Trends Cogn. Sci. 2005, 9 (12), рр. 566-571.
43. Cubo Е. Martin P. M., Martin-Gonzalez J. A., Rodriguez-Blazquez C., Kulisevsky J. Motor laterality asymmetry and nonmotor symptoms in Parkinson’s. Movement disorders. 2010, 25(1), рр. 70-75.
44. Francis A. N., Bhojraj T. S., Prasad K. M., Kulkarni S., Montrose D. M., Eack S. M., Keshavan M. S. Abnormalities of the corpus callosum in non-psychotic high-risk offspring of schizophrenia patients. Psychiatry Res. 2011, 191 (1), рр. 9-15.
45. Haier R. J., Jung R. E., Yeo R. A. Head K., Alkire M. T. The neuroanatomy of general intelligence: sex matters. Neuroimage. 2005, 25 (1), рр. 320-327.
46. Hommet C., Destrieux C., Constans T., Berrut G. Aging and hemispheric cerebral lateralization. Psychol. Neuropsychiatr. Vieil. 2008, 6 (1), рр. 49-56.
47. Logue D. D., Logue R. T., Kaufmann W. E. Belcher H. M. Psychiatric disorders and left-handedness in children living in an urban environment. Laterality. 2015, 20 (2), рр. 249-256.
48. Luders E., Narr K. L., Zaidel E., Thompson P. M., Toga A. W. Gender effects on callosal thickness in scaled and unscaled space. Neuroreport. 2006, 179 (11), рр. 1 103-1 106.
49. Preti A., Vellante M. Creativity and psychopathology: higher rates of psychosis proneness and nonright-handedness among creative artists compared to same age and gender peers. J. Nerv. Ment. Dis. 2007, 195 (10), рр. 837-845.
50. Villarreal G., Hamilton D. A., Graham D. P., Driscoll I., Qualls C., Petropoulos H., Brooks W. M. Reduced area of the corpus callosum in posttraumatic stress disorder. Psychiatry Res. 2004, 131 (3), рр. 227-235.
Контактная информация:
Макарова Ирина Илларионовна — доктор медицинских наук, профессор, зав. кафедрой физиологии ГБОУ ВПО «Тверская государственная медицинская академия» Минздрава России
170100, г. Тверь, ул. Советская, д. 4
Тел. 8(4822) 34-46-95
E-mail: [email protected]
проблемы обучения в норме и патологии
Проблема межполушарных асимметрий одна из самых актуальных проблем современности. Изучение асимметрии полушарий играет большую роль в решении проблемы в образовании. Это связано с тем, что в образовательных учреждениях важно учитывать индивидуальные особенности каждого ребёнка, в том числе и доминирующее полушарие. Различные науки современного мира изучают головной мозг и проникают в проблему межполушарной асимметрии мозга.
У каждого человека можно выделить доминантное полушарие. Выявление доминантного полушария важно знать каждому человеку, потому что это в дальнейшем повлияет на его социализацию, обучение, развитие и поможет разобраться в его способностях. Знания доминирования полушария очень важно и в психологическом плане. Ориентируясь на доминирующие полушарие психолог, логопед и педагог могут использовать специальные методы для коррекции или обучения ребёнка, которые помогут ребёнку легче справиться с заданием [7].
С давних времен люди чётко очерчивали круг «своих» и «чужих». Руководствуясь заложенной в психике человека доли стадности, люди во все времена соединялись в группы, объединения, общества. Поначалу это были совсем примитивные признаки, например, принадлежность человека к определённому роду. Но время шло и по мере развития общества и самого человека, признаки становились всё более изощрёнными.
Человек с правополушарной асимметрией мозга, то есть левша, не может остаться не замеченным в обществе правшей. Он воспринимает информацию, мыслит, выполняет бытовые действия совсем иначе, чем левополушарные или праворукие, что и выдаёт его особенную организацию психики. Совокупность показателей функциональной, сенсорной и моторной асимметрий даёт полную картину в определении индивидуального латерального профиля конкретно взятого индивида. Исходя из этого в современном научном обществе выделяют 3 типа организации головного мозга: левополушарный, правополушарный и равнополушарный типы [1]. Подробно хотелось бы остановиться именно на первых двух, как наиболее распространённых типах организации мозга.
По статистическим данным, доля населения с правополушарной асимметрией мозга не так уж и малочисленна и составляет около 15% населения Земли. Но, несмотря на это левши всегда чувствуют своеобразное давление со стороны общества. В детстве многие родители рьяно пытаются, во что бы то ни стало переучить «несчастного» малыша, сделав из него «нормального» правшу. Когда ребёнок вырастет, он постоянно будет ощущать заинтересованные взгляды в свою сторону, а иногда и достаточно негативное отношение к своей особенности психики. К сожалению, в силу не достаточной проинформированности граждан, порой возникают абсолютно нелепые представления о левшах как о «чужих» нашему социуму, некоторые и вовсе приписывают им психические отклонения. На самом же деле правополушарная асимметрия головного мозга, конечно же, не означает обязательное присутствие у индивида отклонений от нормы развития, а лишь создаёт различия познавательных процессов левшей и правшей [5].
У человека с левополушарным типом организации мозга в качестве доминирующего полушария выступает левое полушарие. Такой индивид имеет склонность к обобщению и абстрагированию, а познавательные процессы носят преимущественно словесно-логический характер. Именно левое полушарие предназначено для проведения операций с символами, условными знаками и словами, что даёт возможность сделать вывод о наличии концептуального и абстрактного типов мышления. Особенностью мышления у правшей служит и наличие так называемых элементов однозначного контекста. Левополушарные индивиды при организации материала знакового характера создают строго однозначно понимаемый текст, без возможности его интерпретации и объяснения иными путями. Иными словами, можно сделать вывод о достаточно педантичном и однобоком подходе к пониманию высказываний в процессе взаимодействия в социуме. К явным преимуществам правшей можно отнести их заострённое чувство долга перед данным ими обещанием, они исполнительны, ответственны, рассудительны, рациональны и принципиальны. Так же для индивидов левополушарного типа характерен большой объём памяти, а речь таких людей лишена аграмматизмов. Но также в их характере присутствуют такие особенности как: недостаточное проявление гибкости, спонтанности, непосредственности. Они привыкли держать всё в себе, не выпуская своих истинных переживаний и эмоций наружу, поэтому могут показаться несколько скрытными. Исходя из структуры левополушарной асимметрии мозга, правшам подойдут такие профессии как: лингвист, инженер, философ, математик. Также они могут являться не только представителями теоретических дисциплин, но и административных должностей [2].
Люди с правополушарным типом организации головного мозга также имеют свои характерные черты. К таким мы можем отнести, к примеру, уже не словесно-логический характер познавательных процессов, а конкретно-образный. Кроме того, левши характеризуются наличием дивергентного типа мышления. Такие люди, как правило, мыслят не стандартно, для них не существует рамок и ограничений, они видят в решении любой проблемы множество различных вариантов и путей их решения, в отличие от узко мыслящих правшей. Правое полушарие мозга по своим функциям отвечает за ориентацию в пространстве, операции с образами реальных и конкретных предметов, а также легко воспринимает пространственные связи между предметами. Левши имеют наглядно-образное мышление, что помогает им видеть ситуацию в целом и прогнозировать изменения, к которым приведёт то или иное действие по отношению к данной ситуации. Правое полушарие мозга, непрерывно регулируя подсознательные процессы и контроль над непроизвольным поведением, производит пространственные и структурированные преобразования информации, даёт оценку сложности и симметрии предмета. Правополушарный тип организации мозга даёт индивиду возможность как бы смотреть вглубь происходящего, помогая видеть не только сам предмет перед собой, но и фон за ним, иметь дело не с центром, а с периферией. Таким образом, правое полушарие обеспечивает равномерное распределение внимания, а не его фокусировку и концентрацию. Являясь органом человеческого бессознательного, правое полушарие даёт возможность одновременного распознавания и анализа большого числа связей, иногда и достаточно противоречивых с логической точки зрения. Люди с правополушарной асимметрией пользуются в жизни скорее интуитивным чутьём и эмоциональными порывами, нежели строгими и логическими доводами. Преимущество данного типа мышления проявляется в ситуациях, которые связаны со сложной информацией, имеющей большое число внутренних противоречий и которую нельзя на первый взгляд свести к однозначному смыслу. Ситуации подобного характера — творческий процесс. Левши, как правило, имеют довольно экспрессивно окрашенную речь, богатую жестикуляциями и интонациями. Таких людей точно не назовёшь нудными и монотонно говорящими. Несмотря на то, что в их речи нет выстроенности, возможна сбивчивость, запинки, а также лишние слова и звуки, левши имеют эмоциональную и богатую речь. Именно про данную категорию людей с восхищением говорят: «А у него язык подвешен!». Левши являются целостными, творческими натурами, способными к открытому выражению чувств. Они открыты миру, наивны и доверчивы, контактны и общительны. Для правополушарного типа организации также характерна способность тонко чувствовать и тяжело переживать сложные жизненные ситуации, легко огорчаться и плакать, либо приходить в гнев, то есть действовать по настроению. Именно поэтому среди левшей можно встретить большое число журналистов, литераторов, организаторов и деятелей культуры [8].
Выделив и проанализировав особенности психики и характера познавательных процессов лево- и правополушарных людей, мы можем сделать вывод о необходимости разных подходов в процессе освоения ими школьной программы. Любой процесс обучения начинается с правильно выстроенной мотивации ребёнка на получение знаний и собственно дальнейшее обучение. Важным компонентом в достижении этого является заинтересованность ребёнка, создание таких условий, которые продемонстрируют ему, что образовательный процесс не является рутинным и скучным явлением. Так, зная особенности мышления правополушарных детей, мы будем делать акцент в их обучении на эстетическую сторону предметов. Для левшей велика потребность в самореализации, получении наивысшего балла и похвалы учителя. Для левополушарного же типа мышления наоборот характерна ориентация на познавательную сторону образовательного процесса. Их привлекает не похвала, а непосредственно само приобретение новых знаний и их глубина. Сами школьные занятия рассматриваются правшами как способ самосовершенствования, развития волевых качеств и мышления. Для них важна не оценка учителя, а сам результат обучения. Кроме того, в процессе усвоения школьной программы важно разделять детей и по их сенсорным особенностям восприятия. Так, можно провести простой тест для определения данного факта. Нужно проследить в какую сторону отводит глаза ребёнок в процессе мышления. Так как познавательная активность первично возникает в одном из полушарий головного мозга, запуская движение глаз, можно предположить, что левополушарные дети будут отводить глаза вправо, а правополушарные – влево. Для левополушарных детей значимой для восприятия будет являться правая полусфера пространства класса, поэтому необходимо посадить такого ребёнка таким образом, чтобы доска и учитель были от него с правой стороны. Для правополушарных детей будет действовать по аналогии тот же принцип, но с заменой стороны полусферы класса [9].
Важной особенностью является и процесс переработки информации во время урока, который у лево- и правополушарных детей происходит по-разному. Левополушарных учеников в классе часто могут называть «тугодумами», из-за их медленного процесса обработки информации. Они медленно и вдумчиво смакуют каждую деталь, для них не характерна работа по принципу «вопрос-мгновенный ответ». Правополушарные же дети наоборот перерабатывают информацию практически мгновенно, но спонтанно и хаотично. Таких учеников учителя называют «торопыжками».
Речевой аспект образовательного процесса выявляет некоторые проблемы в использовании речи право- и левополушарными детьми. Для учеников с доминирующим правым полушарием характерен слабый уровень самоконтроля в выполнении действий. Именно поэтому у правополушарных учеников слабо развит контроль над своей произносительной речью, возможны пропуски слов и проблемы в подборе слов и грамматике. У детей с доминирующим левым полушарием напротив развит контроль над произносительной стороной речи. Их слабой стороной является подведение итогов. Также проблемы могут возникнуть с развитием беглой устной речи, возможна необходимость оказания помощи и в развитии скорописи у ребёнка. Успешно даётся им применение правил и правильное употребление слов в предложении [1].
Существенные отличия типов организации головного мозга мы можем наблюдать и на примере процесса памяти. Для правополушарных учеников с визуальной памятью характерной особенностью является визуализация информации. Такие дети видят слова, будто глазами мозга. У детей правополушарного типа присутствует феномен «врождённой грамотности». Их пальцы и глаза, словно сами пишут за них, зная правильность написания того или иного слова. Но, к сожалению, в процессе обучения в школе «врождённая грамотность» у данной категории детей разрушается. Это происходит по причине применения в программе общеобразовательных учреждений аналитического подхода к изучению грамотности, который не свойственен правополушарным ученикам. Ребёнка с доминирующим левым полушарием можно сравнить с магнитофоном. За счёт присутствующего у ученика аудиального типа памяти, он после получения вопроса как бы прокручивает у себя в голове нужную кассету. А затем ребёнок выбирает из неё кусочек, содержащий правильный ответ. Левополушарный ученик в процессе образовательного процесса зачастую сталкивается с проблемой необходимости трансляции запомнившейся информации в другую модальность. Это происходит из-за склонности учителей к смене модальности в процессе преподнесения материала урока и проведения экзамена. В противовес данной проблеме, левополушарные дети отлично пользуются характерными особенностями своей произвольной памяти. В отличии от правополушарных учеников, которые запоминают в практической деятельности, а не усилием воли, левополушарные дети склонны к зазубриванию. Для запоминания им необходимо многократное повторение и заучивание, которое и используется в школьной программе [7].
Важным является и верно выбранные приёмы и методы в процессе школьного обучения. Для этого необходимо учитывать особенности мыслительных процессов, характерные для детей с разными типами функциональной асимметрии головного мозга. Так, важно помнить о необходимости различных подходов и в процессе обучения с использованием теоретических материалов. Чтение и оценивание слов в тексте левополушарными детьми сравнимо с «атакой». Именно поэтому таким ученикам важно применение именно дискретного подхода в обучении, а то есть индуктивный путь «от части — к целому». К правополушарным ученикам же напротив необходимо применение дедуктивного пути обучения «от общего — к частному». Именно по этой причине часто наблюдаются проблемы усвоения школьной программы правополушарными детьми по методике Зайцева или системе Занкова, так как они предназначены для левополушарного типа организации мозга [8].
Особенности индивидуального латерального профиля ребёнка зачастую определяют и наличие у него возрастных и половых проявлений организации мозга. Если же по каким-либо причинам учителя и родители не учитывают данные особенности функциональной организации мозга, то это может привести к появлению таких проблем как гиперактивность, школьная неуспеваемость и недостатки речевой функции. Последняя является одной из важнейших проблем, так как недоразвитие, задержка или полное отсутствие речи затруднит школьное обучение и будущую социализацию ребёнка.
При работе с детьми с ОВЗ, и особенно с нарушением слуха и речи одной из важнейших проблем является диагностическая работа с ребёнком по прогнозированию особенностей его речевого развития. Решение этой проблемы кроется в анализе профиля функциональной асимметрии [4].
Исследования, проводимые с детьми с общим недоразвитием речи, открывают множество фактов о взаимосвязи индивидуального латерального профиля ребёнка с речевыми нарушениями. Так, именно среди детей с отклонениями, наиболее часто встречаемым является левый профиль функциональной сенсомоторной асимметрии, нежели правый. Также любопытным фактом является то, что при анализе детей из группы с выраженными нарушениями речи мальчики в большинстве своём имеют смешанный тип функциональной сенсомоторной асимметрии, а у девочек же отмечаются левый либо симметричный профили. Если же подвергать анализу интеллектуальные способности детей с ОНР, то мы придём к любопытному выводу. Дело в том, что самые высокие показатели интеллекта наблюдаются в группе нормативно развивающихся мальчиков, а также – мальчиков, имеющих незначительные нарушения речи. Но при этом у детей из обеих групп обязательно присутствует левый тип функциональной асимметрии. Если же брать группу дошкольников с более выраженными речевыми нарушениями, то там мы будем наблюдать наличие самых низких интеллектуальных показателей у мальчиков с правым и смешанным типами функциональной асимметрии. Кроме того, результатами исследования было выявлено, что в группах дошкольников с менее выраженными речевыми нарушениями встречается примерно одинаковое число детей с левым, правым или симметричным профилем. Но, если анализировать группу дошкольников с выраженными проблемами речевого развития, то там мы увидим наибольшее число детей именно с левым профилем функциональной сенсомоторной асимметрии. Исходя из приведённых фактов, мы можем предположить, что наличие левопрофильности у дошкольников с выраженными речевыми нарушениями свидетельствует о паранатальных воздействиях на ребёнка, которые могли привести к снижению уровня активности правого полушария [6].
Особенности индивидуального латерального профиля, кроме речевого аспекта, накладывают отпечаток на интеллект и психическое здоровье ребёнка. Так, по данным исследования, проводимого с детьми с нормой развития, ЗПР и умственной отсталостью, мы можем сделать вывод о специфике проявления асимметрии мозга. По результатам процентного соотношения, мы видим, что процент «чистых правшей» и праворуких в группах детей с ЗПР выше на 1,6-1,8 % по сравнению с группой испытуемых с нормой развития. Но, несмотря на это, в общем соотношении, процент «чистых правшей» и праворуких детей в группах детей с ЗПР и умственной отсталостью самый низкий по сравнению с группой с нормой развития, и составляет от 1,8% до 4 % [3].
Если сравнить левополушарных и правополушарных людей, то можно сделать вывод о том, что эти люди по-разному приспосабливаются к миру, используя свои способности. Доминирующее полушарие поможет человеку определиться со своей будущей профессией, сделает его трудовую деятельность менее сложной. Но также человек должен развивать и то полушарие, которое не преобладает в его жизнедеятельности, ведь оно поможет ему в различных жизненных ситуациях и сделает человека более уверенным в себе [5].
Доминирующее полушарие важно учитывать и в особенности развития детей. Ведь от этого зависит их обучение и воспитание. Можно подобрать различные методы и способы, учитывая доминирующее полушарие, которые помогут ребёнку в дальнейшем развитии и обучение в детском саду и в школе. Как известно, школьные методики обучение главным образом тренируют и развивают левое полушарие, игнорируя развитие правого полушария. С помощью дифференцированного обучения, исследуя специфику когнитивного развития и восприятия ребёнка, можно разработать педагогические методы и способы, учитывая индивидуальные и психофизиологические особенности детей.
Таким образом, мы можем сделать вывод о необходимости своевременного определения индивидуального латерального функционального сенсомоторного профиля ребёнка для его дальнейшего успешного обучения в школе. Ведь, к каждому ребёнку нужен сугубо индивидуальный подход, способный в будущем благотворно сказаться на его психическое и умственное развитие.
Функциональные Ассиметрии Человека / Глава 3. Современное состояние вопроса об асимметриях полушарий мозга
Цель этой главы не столько в
обобщении, сколько в иллюстрации многочисленности направлений исследования
асимметрии полушарий и мозга по разным признакам. С давних пор отмечаются
различия в их строении.
По отношению ко всем частям тела В. В. Бунак отмечал:
«каждый индивидуум представляет собой как бы два рано дифференцировавшихся по
продольной оси полуиндивидуума, правого и левого, развивающихся совместно»
(1926). Полушария мозга симметричны по одним, асимметричны по другим
показателям. Морфологические асимметрии рассматриваются как вторичные,
связанные с более высокой дифференцировкой первично-симметричных закладок
[Пинес Л. Я., 1934], возрастают в филогенетическом ряду, более рельефно
выражены у человека; чем у антропоидов, максимально выражены в неокортикальных
структурах: нижнетеменной, нижнелобной, верхневисочной. В них вариабельны
цитоархитектоническое строение, формы клеток, типы дендритов [Блинков С. М.,
1955; Шевченко Ю. Г., 1972]. Так, поле 42 по размерам больше в доминантном но
речи полушарии, а поле 22/38 — в верхневисочной подобласти субдоминантного
[Блинков С. М., 1955]. Самый большой коэффициент асимметрии отмечен в зонах
Брока и Вернике, получающих максимальное развитие в онтогенезе [Блинков С. М.,
Глезер М. И., 1964]. Как о наиболее ярких морфологических различиях говорят о
преобладании левой височной площадки по сравнению с правой [Geschwind N., Levitsky W., 1968; Galaburda A. et al., 1978; Geschwind N., 1978, и др.].
С помощью количественных методов выявлены достоверные
различия речедвигательных полей 44 и 45 в правом и левом полушариях [Боголепова
И. Н., 1982; Боголепова И. Н. и др. 1983]. Дивергентные архитектонические
различия отмечены в нижней теменной дольке, функционально связанной с
полимодальной сенсорной интеграцией [Eidelberg D. et al., 1984].
Показаны нарастание асимметрий проводящих путей в
филогенетически новых отделах мозга [Дзугаева С. Б., 1965], значение
коллозальной системы [Бианки В. Л., 1985], активное участие комиссур в
межполушарном обмене сенсорной информацией различной модальности, что лежит в
«основе дубликации следов памяти» [Мосидзе В. М., 1985]. Есть указание на то,
что у леворуких и амбидекстров мозолистое тело больше, чем у праворуких [Witelson S., 1985]
и это предполагается связанным с большим двуполушарным представительством
когнитивных функций.
Обсуждаются соотношения между морфологическими и
функциональными асимметриями полушарий мозга. О. С. Адрианов (1985) говорит о
«структурных основах функциональной специализации полушарий мозга», считает
перспективным изучение этих основ, не разделяя при этом «пессимизм» С.
Спрингера и Г. Дейча, заключивших, что неизвестно, действительно ли анатомические
асимметрии «являются морфологической основой функциональной асимметрии между
полушариями» (1983)
Интенсивно развиваются представления о различиях
полушарий в электрической активности. Ее показатели изменяются соответственно
тому, в каком психическом, в частности эмоциональном, состоянии находится
сейчас испытуемый — здоровый и больной [Костандов Э. А., 1977, 1978; Костандов
Э. А., Арзуманов Ю. Л., 1980; Костандов Э. А., Арзуманов Ю. Л., и др., 1981;
Арзуманов Ю. Л., 1985; Мадорский С. В., 1985, и др.].
Вопрос о соотношении различий полушарий по показателям
электрической активности с одной стороны, и по их функциям в деле обеспечения
целостной нервно-психической деятельности — с другой, представляется сложным.
С. Спрингер и Г. Дейч (1983) считают преждевременным «пропагандировать
использование регистрации ЭЭГ и ВП в качестве безоговорочных критериев
межполушарной асимметрии». Но все же изучение электрической активности правого
и левого полушарий в момент покоя и выполнения испытуемым психической деятельности
разного содержания ценно тем, что оно, безусловно, дополняет знания об
асимметрии мозга важными данными.
Исследуются различия мозгового кровотока у бодрствующего
человека в момент выполнения им различных видов психической деятельности
[Шахнович А. Р., Разумовский А. Н., Мякота А. Е., 1976; Lassen N., Inquar D., 1972; Inguar D., 1985, и др.]. Исследовался
регионарный кровоток в двух полушариях одновременно у праворуких мужчин во
время выполнения двух видов деятельности — вербальной и перцептивной. Во второй
задаче испытуемые, рассматривая картинки, содержащие отдельные фрагменты,
должны были сказать, что на них нарисовано. Обнаружились небольшие, но
достоверные различия кровотока: как и ожидалось, средняя величина кровотока при
первой деятельности была больше в левом, при второй — в правом полушарии [Risberg J. et al., 1975].
В нашей работе отмечено увеличение мозгового кровотока в
правом полушарии у больных с поражением и правого и левого полушарий, если они
прослушивали музыкальное произведение; при этом изменения кровотока в левом
полушарии были вариабельны — кровоток уменьшался, оставался на прежнем уровне и
у единичных больных увеличивался [Гасанов Я. К., Брагина Н. Н., Доброхотова Т.
А., Корниенко В. Н., Репин В. Я., 1982].
Авторы всех исследований констатируют, что кровоток в
правом и левом полушариях мозга более сходен, чем различен; это обстоятельство
сохраняется и в том случае, если испытуемый в эксперименте занят деятельностью,
основанной на речи.
За последние десятилетия резко усилился интерес исследователей
к тому, асимметричны ли полушария мозга по химическим характеристикам?
Полученные к настоящему времени данные исследований животных, патологических
состояний (разного происхождения) человека и посмертного изучения его мозга
обобщаются как свидетельствующие о биохимической асимметрии мозга [Луценко В.
К., Курганов М. Ю., 1985].
По величине холинэстеразной активности различаются
анализаторы: симметричные — слуховой, зрительный, кожной чувствительности и
асимметричные — моторный, речевой. Асимметрия холинэстеразной активности
обнаружена только в тех областях коры мозга, пишет В. С. Кононенко (1980),
которым «свойственна функциональная асимметрия» и можно различать правый и
левый тип доминирования холинэстеразной активности.
Показано неравномерное распределение норадреналина в коре
мозга и надпочечниках крыс-самцов (Вистар) и уменьшение этой асимметрии при
развитии экспериментального невроза из-за гипокинезии. В левом и нравом
неокортексе до гипокинезии было соответственно 554±86 и 462±56 нг/г норадреналина,
после 6-недельной гипокинезии — 621±187 и 605±137 нг/г; в левом и правом
гиппокампе до гипокинезии — 641±147 и 1064±216 нг/г, после 6-недельной
гипокинезии — 910±145 и 980±60 нг/г соответственно. P. Poppei и соавт. (1984) привлекают
внимание к резкому уменьшению (исчезновению) асимметрии распределения
норадреналина в исследованных структурах мозга в процессе развития
экспериментального невроза.
Неравномерно распределена гаммааминомасляная кислота
(ГАМК) [Guaneri P. et al., 1985]. Ее содержится больше в ядрах черной субстанции,
переднем четверохолмии правого полушария и вентромедиальном ядре зрительного
бугра, хвостатом ядре левого полушария [Starr M. et al., 1981). Неравно распределены
эндорфины и энкефалины [Hughes J. et al., 1980], специфические рецепторы бензодиазепинов [Robertson H.,
1980], серотонин [Попова Н. К. и др., 1978], оппоидные пептиды и опиатные
рецепторы [Чазов Е. И. и др., 1981; Вартанян А. Г. и др., 1982; Бакалкин Г. Я.
и др., 1984, и др.]. P. Flor-Henry (1986) отмечает, что в левом полушарии
больше дофамина, ГАМК, ацетилхолина, а в правом — серотонина, норадреналина.
Установлена асимметрия в распределении пептидов, участвующих в регуляции
двигательных функций. Г. Н. Крыжановский и соавт. (1984) полагают, что
предпосылки к латерализации формируются на молекулярном уровне.
Неодинакова чувствительность правых и левых подкорковых
структур к действию фармакологических препаратов [Дутов А. Х., Анохов С. С,
1983]. Дискинезии, обусловленные нейролептическими средствами, у человека
проявляются в большей степени в правых конечностях, что свидетельствует о
большей чувствительности к ним допаминэргических систем левого полушария [Waziri R., 1980].
Есть указания на большее воздействие аминазина на структуры левого полушария
мозга [Максимович Я. Б и др., 1985], алкоголя на структуры правого [Костандов
Э. А. и др., 1981; Рещикова Т. Н., 1985].
За изложенными проявлениями химической асимметрии, может
быть, кроется закономерность, подобная сформулированной еще в прошлом веке Л.
Пастером: «…в рассуждения и физиологические исследования проникла идея о
влиянии молекулярной диссимметрии естественных органических соединений,
возникло представление о факторе, имеющем очень большое значение и
обусловливающем в настоящее время, может быть, единственное, отчетливо
выраженное различие, которое мы можем обнаружить между химией неживой природы и
химией живой природы» (1960). Допускаемая закономерность на уровне человека и
формирования его сознания должна, по всей вероятности, проявляться в
соответствии с инвариантным, наличным в мозге всех людей (и животных)
несходством полушарий по форме, пространственной характеристике по правизне и
левизне. Полушария представляют собой как бы зеркальные отражения друг друга.
Проблема установления причин и закономерностей формообразования
у живых организмов вообще признается «одной из наиболее трудных как и
методологическом, так и в экспериментальном отношении» [Преображенский Б. В.,
1983]. Открытые Н. И Вавиловым (1920) законы гомологических рядов
наследственной изменчивости обнаружили порядок (симметрию) и полиморфизме гено-
и фенотипических признаков, близких родов и видов. Изменчивости форм,
охватывающей практически все основные «геометрические фигуры», сопутствовали
химические и физиологические особенности. «Близкие виды растений
характеризуются сходством химического состава, выработкой близких или одних и
тех же специфических химических соединений» [Вавилов Н. П., 1965]. По Н. И.
Вавилову, возможность появления новых признаков, морфологических или
биохимических, имеет свои границы и открывает путь для прогнозирования еще
неизвестных структурно-функциональных вариантов. Учение о гомологии в настоящее
время широко используется для характеристики структурно-функциональных
отношений и позволяет найти общий план строения биологических объектов с низком
симметрией [Шафрановский И. И., 1971].
Поиск каких-либо внутренних различий правых и левых форм
биологических объектов оказался безуспешным. Изучали воздействия внешней среды,
например, гелиотропического фактора на правизну — левизну формы объектов
[Смирнов Л. Л., 1950; Дубров А. П., 1987], на право-левостороннюю ориентировку
спиралевидных структур в клетках [Алпатов В. В., 1951].
Обобщая фактический материал о полиизомерии биологических
объектов, В. Б. Касинов (1973) приходит к выводу, что все попытки соотнести
киральность объекта с молекулярной или стереохимической специфичностью
протоплазмы не получили убедительных подтверждений. При этом, по мнению автора,
и генетический контроль киральности низших организмов не является абсолютным, так
как встречаемость правых и левых форм колеблется от равной через преобладание
одного биоизомера до почти полного отсутствия одного из изомеров.
Обосновывается положение о принципиальном сходстве
геометрических форм растений в разных геологических периодах [Мейен С. В.,
1971], о возможности предсказания новых форм растений [Вавилов Н. И., 1965], об
экологической обусловленности форм в биологии, их определяемости образом жизни
[Гиляров М. С, 1944; Беклемишев В. Н., 1964], о приложимости математических законов
к толкованию процессов формообразования [Любищев А. А., 1982].
Содержание понятия формы в последние годы получило более
широкое толкование. Понятие органической формы объединяет всю совокупность
пространственно-временных отношений на макроуровне и глубоких уровнях
организации [Студитский А. Н., 1979; Струков А. П., Хмельницкий О. К., Петленко
В. П., 1983]. Форма связывается с пространственной упорядоченностью частей в
целом, устойчивостью при непрерывной динамике метаболических процессов
[Саркисов Д. С, 1977]. Отступление от абсолютизации формы, понимание ее как
одного из элементов структуры, очевидно, не дает оснований для сопоставления
функций непосредственно с правизной и левизной больших полушарий мозга в отрыве
от материальных процессов, связанных с этой формой.
Из всех асимметрий в нашей работе
главное значение придается правизне — левизне полушарий. Правизна — левизна
отражают, по-видимому, наиболее фундаментальное несходство гемисфер мозга, в
рамках которого проявляются все другие асимметрии полушарий.
Сформулировано множество гипотез об
асимметрии функций полушарий мозга. Различия функций используются иногда в
высказываниях авторов о происхождении функциональной асимметрии мозга.
Так, в происхождении асимметрии мозга предполагаются важными
праворукость и специфически человеческие интеллектуальные процессы [Walker S., 1980],
особое развитие правой руки в силу приспособленности к ней орудий труда [Глумов
Г. М., 1985], левополушарная локализация сознания [Popper K., Eccles J., 1977; Eccles J., 1973], необходимость сегрегации
несовместимых функций, например, анализа и синтеза [Semmes J., 1968; Levy J., 1969], целесообразность
экономного расходования нервной энергии, не дублируя высших функций [Bradschow J.,
1985] и т. д.
Вместе с тем речь идет об огромном
фактическом материале, о множестве уже выявленных асимметрий функций. Знание
этих данных абсолютно необходимо для понимания природы функциональной
асимметрии мозга человека и ее отличий от асимметрии мозга животных.
Получены веские данные в пользу различной обработки
информации двумя полушариями. Широкое распространение получила гипотеза Л. И.
Леушиной, А. А. Невской, М. Б. Павловской (1981, 1985) об обработке зрительной
информации: левое полушарие действует по классификационно-дискриминантному
методу (выработка решающего правила, позволяющего относить изображение к тому
или иному из ожидающихся классов), правое — по структурному методу (описание
иерархической структуры изображения) По мнению авторов, «способ переработки
информации — зрительной и других модальностей является определяющим для
развития речевых функций в одном из полушарий, а именно классификация образов в
левом полушарии, давая более обобщенное и абстрактное отражение объектов
окружающего мира, создает базис формирования речи как высшей формы абстракции».
Для разделения объектов на классы дискриминантный метод проще, он дает короткие
кодовые описания, но может быть применен только к выученному алфавиту
зрительных образов; структурный метод сложнее, но более универсальный и мощный
и может быть использован при описании новых, ранее незнакомых изображений
[Невская А. А., 1985]. Левое полушарие обеспечивает «инвариантное к размеру,
местоположению и, возможно, к повороту описание формы изображений. Эти
схематизированные описания отображены в метрическом пространстве зрительных
образов, в котором возможно установление степени близости и сходства между
ними. Можно думать, что это пространство организовано одинаково для разных лиц»
[Леушина Л. И., Невская Л. Л., Павловская М. Б., 1985].
О зрительной системе говорится как о двух подсистемах,
каждая из которых работает «по своим особым принципам». Одна преимущественно
связана с левым, другая — с правым полушарием. «Наличие в мозге двух таких
подсистем позволяет независимо решать ряд важных, но несовместимых на одном
нейронном субстрате задач». Полноценное узнавание возможно лишь при совместной
работе обоих полушарий мозга. Левое полушарие выполняет схематическое (с
точностью до класса) распознавание отдельных объектов. В правом полушарии
находится основная зрительная память с «записанными» для каждого класса
объектов реализациями (изображениями конкретно виденных представителей данного
класса). Сведения о классе распознанного объекта передаются в правое полушарие,
что сокращает поиск в зрительной памяти и позволяет довести распознавание до
уровня конкретного идентификации [Левашов О. В., 1985].
Долговременная память, помехоустойчивость зрительного
восприятия и возможность компенсации имеющихся расстройств больше страдают при
резекции правого гиппокампа [Меерсон Я. А., 1982].
В левом полушарии происходит высший семантический анализ
и осознание раздражителя. Оно доминирует при восприятии осознаваемой
эмоционально отрицательной информации; в правом производится
зрительно-пространственный анализ осознаваемых вербальных и невербальных
раздражителен, и результаты передаются в левое полушарие», организуются
безотчетные эмоции [Костандов Э, Д., Арзуманов Ю. Л., 1980; Костандов Э. А.,
1983].
Левее полушарие работает как планирующий, аналитический,
последовательный процессор или препозиционно: оперирует дискретными понятиями,
соответствующими целым классам объектов, устанавливая отношения между ними.
Правое полушарие работает аппозиционно: обеспечивает целостное синтетическое, аналоговое
описание мира [Bogen J., 1969, 1975].
Полушария мозга асимметричны в восприятии и обозначении
цветов. Правое обеспечивает словесное кодирование основных цветов с помощью
простых высокочастотных названий (красный, синий). Здесь характерны минимальные
латентные периоды называния и точное соответствие названий физическим
характеристикам основных цветов. «Правополушарный язык обозначения
промежуточных цветов беден»; в нем редко появляются предметно соотнесенные
названия, но они точно соответствуют цвету предмета. В целом правое полушарие
ответственно за формирование жестких связей между предметом и цветом, цветом и
словом, словом и сложным цветным образом предметного мира. Левое полушарие
обеспечивает словесное кодирование цветов с помощью относительно редких в
языке, специальных и предметно соотнесенных названий. При угнетении левого
полушария из лексикона исчезают такие названия промежуточных цветов, как
оранжевый, терракотовый, вишневый, цвет морской волны. В целом левое полушарие
не фиксирует жесткие связи между предметом и цветом, цветом и словом, словом и
сложным цветным образом. В основе деятельности левого полушария лежат широкие
ассоциативные связи понятия со словом, относительно оторванные от
конкретно-чувственного анализа предметного мира. Эти данные получены при
изучении больных, подвергавшихся унилатеральному электросудорожному лечению с
функциональным угнетением правого или левого полушария. Н. Н. Николаенко (1985)
на основе этих данных говорит о существовании двух систем узнавания окраски
объектов. Система правого полушария ответственна за «изоморфное чувственное
отражение качества предмета», левое — за понятийное его отражение.
У тех же больных (после право- или левостороннего
электросудорожного припадка) изучались последовательность выбора цветов,
группировка цветов, положение промежуточных цветов в цветовом пространстве и
называние промежуточных цветов [Николаенко Н. Н., Родионов В. Д., 1985]. До
припадка больные в первую очередь отбирают зеленые, затем красные, пурпурные,
оранжевые и наконец — синие цвета: «последовательность выбора цвета имеет
направление от средневолновой (зеленой) части спектра к длинноволновой
(красной) и затем к коротковолновой (синей)». При угнетении правого полушария
сохраняется эта последовательность, левого — изменяется: от длинноволновой
части спектра (красные, оранжевые, желтые цвета) — к средневолновой (зеленый) и
далее к коротковолновой (синий цвет) части. В этой работе авторами используются
понятия «объективное цветовое пространство» и «перцептивное цветовое
пространство». отличающееся от первого «неравномерностью». «Перцептивное
цветовое пространство, организуемое правым полушарием, представляет собой
пространственно строго упорядоченную систему, изоморфную видимому спектру».
Пространственная правильность передачи видимого спектра опосредуется формирующимся
у человека внутренним образом цветового пространства, которое имеет свою
систему координат. Тот факт, что правое полушарие, как пишут авторы,
«предпочитает» выбирать в первую очередь красные цвета, позволяет предположить,
что начальной точкой отсчета в системе координат перцептивного цветового
пространства является длинноволновый (красный) участок спектра. Это
пространство представляется авторами как совокупность относительно
изолированных структурных образований фокусных цветов. «В цветовом пространстве,
организуемом левым полушарием, признаки конкретного цвета (насыщенность,
светлота) не выделяются, исчезает пространственная правильность передачи
спектра, т. е. утрачивается изоморфное отображение физической картины мира
цветов». Это цветовое пространство имеет укрупненный и высоковзаимосвязанный
характер, что, по предположению авторов, служит основой обобщенно-абстрактного
или категориального отношения к цвету, основой для формулирования понятий.
Суждения авторов можно понять так, что у полушарий мозга различны «перцептивные
цветовые пространства» или полушария мозга асимметричны по характеру «перцептивного
цветового пространства».
Имеются данные о различии функций полушарий мозга в
формировании слухового восприятия. Показана специализация правого полушария
мозга к опознанию пространственных характеристик стимула. При его Поражении
«выявляется смещение четкой и стабильной в норме внутренней системы координат,
отражающей» экстраперсональное пространство. Говорится о «наличии модели
внешнего сенсорного пространства в структурах мозга». Такая модель, по
предположению авторов, должна располагаться в упомянутой системе координат, и
«первоочередными по важности структурами такой модели внешнего пространства»
являются «классические центры сенсорных систем», именно в них происходит тонкая
обработка данных о пространственных характеристиках стимула. Необходим, по
мнению Я. А. Альтмана, С. Ф. Вайтулевича, С. П. Порка (1185), еще определенный
и четкий уровень отсчета, на который необходимо наложить внешнее сенсорное
пространство. Авторам представляется вероятным, что таким уровнем отсчета
является схема тела, присутствующая в разных структурах мозга.
Главное направление исследований асимметрии мозга
началось более 100 лет тому назад на основе изучения нарушений речи при
очаговых поражениях левого полушария [Broca P., 1865; Wernike C., 1874], затем —
у больных после расщепления мозга [Sperry R., Gazzaniga M., 1967; Nebes B.,
Sperry R., 1971; Gazzaniga M., 1974; Zaidel E., 1978, и др.]. В последнее время
интересные данные получены в ходе изучения больных после» право- и
левостороннего электросудорожного припадка [Балонов Л. Я., Деглин В. Л., 1976;
Балонов Л. Я., Баркан Д. В. и др., 1979; Балонов Л. Я., Деглин В. Л.,
Черниговская Т. В., 1985, и др.]. Современные представления отличаются от
прежде сложившихся: функции полушарий мозга неравный в формировании не только
фонетического, но и морфологического, лексического, синтаксического,
семантического уровней языка. Участие в организации речевой деятельности и
правого и левого полушарий «осуществляется при постоянном и гибком их
взаимодействии, обеспечивает возможность двоякого познания внеречевой
действительности, создавая целостный чувственный-иконический образ мира и
проверяя его истинность через построение рациональных логических моделей»
[Балонов Л. Я., Деглин В. Л., Черниговская Т. В., 1985].
Д. А. Кауфман, О. П. Траченко (1985) показали, что
преимущественное участие левого полушария необходимо в случае повышения
требований к точности фонемной идентификации, например, при распознавании
логотомов (бессмысленных слогосочетаний). которые анализируются как ряд
конкретных дискретных фонематических единиц. Доминирование левого полушария
выявляется при опознании лексем, играющих особо важную роль в синтаксическом
оформлении высказываний (служебные слова), в случае вторичного происхождения
лексем от слов другого класса (относительные прилагательные) или принадлежности
слова к позднейшим слоям лексики (жаргонные глаголы). Преимущественная роль
левого полушария выявляется также при опознании абстрактных слов, т. е. слов, в
значении которых малая степень образности сочетается с широкой полисемией.
Преимущество правого полушария или возможность опознания как в левом, так и в
правом полушариях установлены для знаменательных слов, что определяется большей
степенью их конкретности и образности, при узком наборе стоящих за ними
значений. По мнению авторов, «…результаты, свидетельствующие о латерализации
восприятия разных типов вербальных стимулов, имеют ограниченное значение в том
смысле, что они не могут рассматриваться как аргумент в пользу представления о
жесткой привязанности восприятия той или иной категории слов к одному из
полушарий. Выводы исследования могут распространяться только на условия
опознания нулевых форм лексем, предъявлявшихся вне контекста. Вполне возможно,
что в условиях естественной коммуникации латерализация восприятия тех же
лексем, входящих в состав связного высказывания, может быть совсем иной и
зависеть от особенностей той синтаксической структуры, в конструкцию которой
они встроены».
На основе анализа различий распада и последующего
восстановления разных составляющих целостной психики после угнетения правого
или левого полушария мозга В. Л. Деглин (1984) формулирует гипотезу о
семиотической природе функциональной асимметрии мозга, согласно которой
последняя выявляется только при обработке знаковой информации и отсутствует при
обработке информации, не имеющей знакового характера. Ведущим, по автору,
является принцип семиотического дублирования: объективная реальность получает в
сознании двойное отражение; мозг параллельно создает две знакомые модели мира:
правое полушарие — иконическую, левое — символическую. «Семиотическая
формулировка различий функции правого и левого полушарий позволяет объединить и
уточнить другие, ранее предложенные формулировки».
Данные изучения асимметрии функций мозга животных в
сравнении с результатами исследований асимметрии мозга человека легли в основу
индуктивно-дедуктивной гипотезы В. Л. Бианки (1985), согласно которой индукция
преимущественно связана с функционированием левого, дедукция — правого
полушария. По мнению автора, с позиций этой гипотезы можно объяснить все
основные дихотомии (аналитико-холистическую, одновременно-последовательную,
пространственно-временную) относительно частных признаков обработки информации,
которые могут быть рассмотрены как следствия индуктивно-дедуктивной гипотезы.
Подчеркивается взаимодополняющее сотрудничество полушарий
мозга, функциональная асимметрия их предполагается эволюционно связанной с
развитием речи, возникновением нового качества в деятельности высших отделов
головного мозга [Костандов Э. А., 1983].
В основе асимметрии мозга Г. А. Кураев (1983) предполагает
механизм доминанты. Асимметрия является необходимым условием реализации
процессов высшей нервной деятельности человека и животных. Нейрофизиологические
механизмы замыкания временной связи реализуются на основе исходной
межполушарной асимметрии состояния активности заинтересованных структур мозга,
последующего формирования однотипного их состояния и завершаются латерализацией
в одном полушарии процессов, обеспечивающих сигнальную реакцию. Динамика
процессов замыкания временной связи идет от асимметрии к симметрии и вновь к
асимметрии активности полушарий головного мозга.
Перечень высказываний, гипотез
можно бы и продолжить. Но из изложенного видно, что о неравнозначности функций
полушарий мозга можно говорить по отношению к разнообразным составляющим
целостной нервно-психической деятельности. Поскольку представленные гипотезы
довольно многообразны, то, естественно, возникает вопрос: не существует ли
некая единая закономерность, определяющая все проявления функциональной
асимметрии полушарий мозга?
Все последующие главы книги
посвящены по существу попытке обоснования одного из возможных ответов на последний
вопрос. Эта попытка осуществляется на основе анализа данных клинических
исследований авторов.
левый глаз — правое полушарие
Карина Каренина, Андрей Гилёв
«Природа» №12, 2015
Карина Андреевна Каренина — младший научный сотрудник кафедры зоологии позвоночных Санкт-Петербургского государственного университета. Занимается изучением социального поведения животных, асимметрии восприятия сенсорной информации, поиском проявлений межполушарной асимметрии в поведении животных в природе. |
Андрей Николаевич Гилёв — кандидат биологических наук, младший научный сотрудник той же кафедры. Область научных интересов — поведенческая экология, когнитивные способности животных, асимметрия мозга и поведения, эволюция асимметрии в использовании конечностей. |
Один из основных принципов функционирования мозга позвоночных животных — принцип асимметрии. Вклад левого и правого полушарий головного мозга в осуществление различных функций неравнозначен [1]. Подобными предложениями начинается большинство современных научных публикаций, посвященных вопросам асимметрии мозга и поведения. В англоязычной научной литературе такое «введение в тему» носит скорее формальный характер, ведь даже у биологов очень отдаленных специальностей общеизвестность и справедливость подобных утверждений не вызывает сомнений. Однако такое отношение к межполушарной асимметрии сложилось сравнительно недавно. Вплоть до начала XXI в. широко распространено было мнение, что функциональная асимметрия мозга характерна исключительно для человека и, по-видимому, имеет отношение к высокому уровню развития человеческого интеллекта [2].
Первые исследования, доказывающие существование асимметрии в функционировании мозга других видов, помимо человека, относятся к 70–80-м годам XX в. В. Л. Бианки выявил основные особенности работы левого и правого полушарий, а также сделал вывод, что функциональная специализация полушария не абсолютна, т. е. в зависимости от выполняемого процесса доминировать может то одно, то другое полушарие [1, 3]. Было обнаружено, к примеру, что левое полушарие мозга преимущественно контролирует пение у канареек [4] и распознавание пищевых объектов у куриц [5], а правое полушарие доминирует в процессах, обеспечивающих ориентацию в пространстве у крыс [6].
Несмотря на убедительную доказательную базу, первые свидетельства существования межполушарной асимметрии у животных зачастую резко критиковались или даже игнорировались научным миром [2, 7], вероятно, еще неготовым отвергнуть идею об уникальности принципиальных основ функционирования человеческого мозга. Но время расставило все по своим местам. На сегодняшний момент существование асимметрии функционирования нервной системы доказано как экспериментально, так и наблюдениями в природе, на разнообразнейших объектах животного мира — без преувеличения, от мухи до слона. Накопление в течение последних трех десятилетий массива информации о неравноценности функций левой и правой половин мозга наконец-то привело к тому, что фундаментальность и древнее происхождение функциональной асимметрии мозга были признаны широкой научной общественностью. Важно отметить, однако, что в основе современных представлений об асимметрии мозга лежит понимание основного свойства парного мозга — его двусторонней симметричности. Общий план строения мозга подчиняется принципу билатеральной симметрии, согласно которому одна половина зеркально повторяет другую. Однако детальное рассмотрение выявляет относительность принципа симметрии как в строении, так и в функционировании мозга. На основе парных, относительно симметричных структур возникает выраженная функциональная асимметрия полушарий мозга [1, 3, 6].
«Правый» и «левый» мир
Базовые различия между полушариями мозга, возникшие, как предполагают, на ранних этапах эволюции позвоночных [8], могут проявляться в поведении животных по-разному. К примеру, для многих видов позвоночных животных характерна моторная латерализация — неравное участие правой и левой половин тела в движении, связанное с асимметрией функционирования нервной системы [9]. Такая латерализация может выражаться в асимметрии поворотов тела и в асимметричном использовании парных конечностей. Другое распространенное проявление межполушарной асимметрии — сенсорная латерализация, т. е. неодинаковая восприимчивость органов чувств на левой и правой сторонах тела к определенным стимулам. Асимметрия восприятия сенсорных стимулов обусловлена специализацией левого и правого полушарий на обработке информации разного типа [10]. В поведении животных сенсорная латерализация может проявляться в виде предпочтения воспринимать определенный стимул левым или правым органом чувств (например, поворачиваться к стимулу одним ухом), а также в более высокой скорости и выраженности реакции на стимул при восприятии его одним из парных органов [11].
Наиболее детально на сегодняшний день изучена латерализация зрительного восприятия. Несмотря на значительные различия в организации зрительной системы, представители разных групп позвоночных по-разному воспринимают и обрабатывают информацию, поступающую от левого и правого глаза. Это явление описано для костных рыб, амфибий, рептилий, птиц и млекопитающих. Зрительная латерализация может проявляться при восприятии различных типов стимулов, например социальных партнеров, пищевых объектов, опасности, пространственных ориентиров [2]. Серия экспериментов на цыплятах домашней курицы служит одним из классических примеров исследований зрительной латерализации. Цыплят выпускали на экспериментальную площадку, по которой были рассыпаны зерна вперемешку с мелкими камешками. В разных вариантах эксперимента цыплята имели возможность использовать оба глаза либо только левый или правый глаз — другой был закрыт непрозрачным колпачком. Оказалось, что, если цыплята могли видеть обоими глазами или только правым, они практически не совершали ошибок, выбирая зерна среди камешков. В то же время, когда цыплята использовали только левый глаз, они с одинаковой частотой клевали зерна и камешки. Так как зрительная информация из правого глаза цыпленка преимущественно поступает в левое полушарие мозга, результаты описанных экспериментов указывают на доминирующую роль левого полушария в распознавании пищевых объектов [12]. Подобная латерализации при восприятии пищи была обнаружена и у представителей других групп позвоночных [13]. Например, костные рыбы и хвостатые амфибии преимущественно реагируют на пищевой стимул, находящийся в поле зрения правого глаза.
Модельными для изучения зрительной латерализации стали животные, у которых глаза расположены на голове латерально, т. е. по бокам. Человек, например, к таким видам не относится, ведь у нашего вида глаза расположены фронтально, т. е. спереди и в одной плоскости. У животных с латеральным расположением глаз поля зрения левого и правого глаза перекрываются лишь незначительно и из-за полного (или практически полного) перекреста зрительных нервов информация из одного глаза поступает преимущественно в противоположное полушарие мозга [14]. У видов с подобным устройством зрительного анализатора мы можем исследовать межполушарную функциональную асимметрию без применения инвазивных методик, просто анализируя поведение животных. Само по себе латеральное расположение глаз не определяет проявление латерализации. Однако многочисленные исследования подтверждают, что предпочтение держать определенные объекты в поле зрения одного из глаз у видов с латерально расположенными глазами обычно отражает доминирующую роль противоположного полушария в обработке информации об объектах такого типа. Предполагается, что части пространства, видимые левым и правым глазом, воспринимаются мозгом неодинаково и во взаимодействии животного с окружающей средой участвуют две разные, но взаимосвязанные системы: «левый глаз — правое полушарие» и «правый глаз — левое полушарие» [15].
Социальное полушарие
Одним из наиболее активно развивающихся направлений в исследованиях асимметрии мозга и поведения стало изучение «социальной латерализации» — асимметричного восприятия особей своего вида. Регулирование социального поведения — одна из наиболее сложных и многокомпонентных функций, которые осуществляет мозг позвоночных животных. В различные аспекты социального поведения могут быть вовлечены процессы, осуществляемые как левым, так и правым полушариями. Однако именно правое полушарие в научной литературе оправданно называют «социальным», ведь оно играет доминирующую роль в контроле большинства социальных реакций у различных видов позвоночных, включая человека [16]. В зрительном узнавании знакомых особей своего вида, к примеру, участвует преимущественно правое полушарие мозга, причем такая асимметрия выявлена как у млекопитающих, так и у птиц [17]. Преимущественная роль правого полушария в проявлении агрессивных реакций, направленных на особей своего вида, известна у амфибий, рептилий, птиц и млекопитающих. Социальная латерализация находит отражение в поведении животных — как в эксперименте, так и в природе. Если малька рыбы поместить в аквариум с зеркальной стенкой, то осматривать «другую особь», т. е. собственное отражение, он будет преимущественно левым глазом (направляющим информацию в правое полушарие). Сходным образом, жеребцы одичавшей домашней лошади во время поединков предпочитают держать противника в поле зрения левого глаза [18].
Асимметрия материнско-детских отношений
Одно из наиболее ярких проявлений асимметрии в социальном поведении человека — предпочтение держать ребенка с левой стороны от себя. Впервые научное описание этого феномена было опубликовано американским психологом Ли Солком в 1960 г. [19]. Он выяснил, что значительное большинство женщин предпочитает держать своего ребенка левой рукой на левой стороне тела. Впоследствии было обнаружено, что такое левостороннее предпочтение существует в различных культурах и проявляется в различных ситуациях: когда ребенка держит на руках его мать или отец, нерожавшая женщина или подросток, а также в случаях, когда вместо живого ребенка женщина держит игрушку, имитирующую младенца. Однако подобная латерализация не уникальна для человека. Так, большинство самок горилл (Gorilla gorilla) и шимпанзе (Pan troglodytes) предпочитают держать детенышей с левой стороны тела [20].
Научные публикации, в которых говорится, что асимметрия в расположении ребенка на руках — не просто любопытный феномен, но и важный критерий физиологического состояния матери, вызвали волну интереса исследователей к этому вопросу в течение последнего десятилетия. Было обнаружено достоверное влияние эмоционального состояния матери на асимметричное расположение ребенка. Более того, оказалось, что женщины, страдающие послеродовой депрессией, не проявляют левостороннего предпочтения [21]. Несмотря на то что асимметрия расположения ребенка относительно матери была обнаружена более полувека назад, механизмы возникновения данного феномена и факторы, определяющие его проявление, остаются не до конца понятными. Существует две основные гипотезы, объясняющие возникновение пространственной асимметрии между матерью и ребенком [19, 21]. Первая связывает данный феномен с моторными предпочтениями, проявляющимися в повседневной жизни человека. Можно предположить, что женщина держит ребенка левой рукой, так как правая должна оставаться свободной для выполнения какой-либо работы. Однако такое предположение не подтверждается данными о практически одинаковом числе матерей, предпочитающих держать ребенка левой рукой, среди правшей и левшей [19]. Вторая гипотеза объясняет асимметрию во взаимном расположении матери и ребенка с точки зрения асимметрии сенсорного восприятия. Предполагается, что левостороннее преобладание возникло из-за предпочтения держать ребенка в периферическом поле зрения левого глаза и, таким образом, направлять поток зрительной информации о нем в первую очередь в правое полушарие мозга [21]. Именно такое положение предположительно обеспечивает наиболее точное и быстрое осознание родителем эмоционального состояния ребенка, так как правое полушарие играет ведущую роль в подобных процессах восприятия социальной информации. Асимметричное расположение относительно родителя может иметь большое значение и для самого ребенка. Было показано, что, когда мать держит ребенка левой рукой, ему лучше видна левая половина лица матери, на которой более интенсивно отражаются эмоции. К тому же наблюдать мать в таком положении ребенок может преимущественно левым глазом, что способствует более эффективному восприятию социальной информации, исходящей от матери. Преимущественное расположение с определенной от нее стороны может оказывать влияние на формирование социальной латерализации. Так, дети, чьи матери держали их правой рукой, во взрослом возрасте проявляют пониженную нормальную межполушарную асимметрию в процессе восприятии лиц [22].
Смотрю левым глазом на себе подобных
У приматов использование передних конечностей — неотъемлемый компонент родительско-детских взаимоотношений, поэтому отдельно оценить вклад моторной и сенсорной латерализаций в возникновение асимметричного расположения особей крайне сложно. Изучение видов, у которых конечности напрямую не задействованы в поддержании близости между матерью и детенышем (мать не удерживает детеныша, а детеныш не держится за мать), позволит с точностью определить, может ли возникнуть асимметрия расположения детеныша лишь под воздействием латерализованного сенсорного восприятия особями друг друга. Это и стало целью нашего исследования. В качестве объектов были выбраны два вида парнокопытных, обитающих на территории России: сайгак (Saiga tatarica tatarica) и северный олень (Rangifer tarandus tarandus). Животные обоих видов обитают на открытых пространствах, имеют хорошо развитое зрение, а их глаза расположены на голове латерально. Кроме того, детеныши выбранных видов с раннего возраста самостоятельно следуют за своими матерями. Перечисленные особенности делают сайгака и северного оленя подходящими объектами для изучения природы асимметрии в расположении матери и детеныша у млекопитающих.
Мы решили исследовать естественное поведение животных в природе, поэтому для полевых работ были организованы экспедиции. Наблюдения за дикими северными оленями проводились в Красноярском крае, в низовьях р. Хеты, а за сайгаками — в Астраханской области, на территории государственного природного заказника «Степной». Используя специальные замаскированные укрытия и маскировочные костюмы, мы проводили фото- и видеосъемку поведения самок с детенышами. При последующем анализе полученного материала мы оценили зрительную латерализацию во время социальных взаимодействий животных.
Перемещаясь вместе с матерью, основную часть времени детеныш проводит от нее сбоку. Объекты нашего исследования относятся к видам с латеральным расположением глаз, поэтому, находясь сбоку от матери, детеныш видит ее преимущественно левым или правым глазом. В ходе наблюдений мы отмечали, какое положение выбирает детеныш, приближаясь к матери сзади. Мы анализировали ситуации, в которых именно он был инициатором объединения пары и выбирал свое положение в пространстве относительно матери. У видов с латерально расположенными глазами предпочтение поворачиваться определенной стороной тела к социальному объекту — свидетельство сенсорной, а не моторной латерализации [14, 17]. Поэтому односторонние предпочтения детенышей при выборе положения относительно матери мы интерпретировали как проявление латерализованного зрительного восприятия.
Результаты нашего исследования показали, что детеныши сайгака и северного оленя предпочитают располагаться в пространстве таким образом, чтобы мать находилась в поле зрения левого глаза. В результате, когда самка с детенышем перемещались совместно, в большинстве случаев он находился справа от нее. Такие предпочтения детеныши проявляли в различных типах активности, например во время отдыха, пастьбы и бегства при опасности. Зрительная латерализация была обнаружена у детенышей разного возраста, от новорожденных до подростков двух-трех месяцев от роду. Наши наблюдения показали, что как у сайгаков, так и у северных оленей в основном детеныши поддерживали пространственную близость с матерью, а не наоборот. Очевидно, что при перемещении пары мать — детеныш основная задача детеныша следовать за матерью, не терять ее из виду. Мать в первую очередь занята выбором направления движения, контролем за безопасностью, взаимодействиями с другими особями своего вида, т. е. ее внимание обращено скорее на окружающую обстановку, чем на детеныша. Несмотря на это, в определенных ситуациях именно мать инициирует сближение и контакт с детенышем. Латерализация в ситуациях, когда мать подходит к своему детенышу и выбирает определенное положение относительно него, — предмет будущих исследований.
Сопоставляя полученные результаты с данными исследований на приматах, можно проследить существование двух вариантов асимметрии в расположении матери и детеныша у млекопитающих. У приматов мать держит детеныша на руках и таким образом определяет его положение в пространстве. Сенсорная латерализация матери приводит к тому, что детеныш преимущественно находится слева от матери. В то же время у исследованных копытных детеныш сам определяет свое положение в пространстве, и его латерализованное поведение приводит к тому, что в большинстве случаев он располагается от матери справа. Несмотря на разное результирующее положение, оба варианта асимметрии между матерями и их детенышами, по всей видимости, обусловлены одной и той же социальной латерализацией, а именно предпочтением воспринимать социальную информацию с помощью системы «левый глаз — правое полушарие» [23]. Когда детеныш сайгака или северного оленя находится справа от матери и держит ее в поле зрения левого глаза, социальная информация от матери поступает преимущественно в его правое полушарие. Это подтверждают исследования организации зрительной системы копытных — около 90% нервных волокон от каждого глаза идут в противоположное полушарие мозга [18]. Таким образом, асимметричное расположение детеныша относительно матери — одно из проявлений правополушарного доминирования в регулировании социального поведения. В отличие от приматов, у исследованных видов передние конечности не участвуют напрямую во взаимном расположении матери и детеныша, поэтому мы можем исключить влияние моторной латерализации [24]. Наши результаты позволили понять, что асимметрия расположения матери относительно детеныша у млекопитающих может возникать вследствие асимметрии зрительного восприятия детенышем матери.
Наблюдая за поведением сайгаков в природе, помимо материнско-детских контактов, мы регистрировали латерализацию в отношениях между двумя детенышами, а также между взрослыми особями. Оказалось, что предпочтение смотреть на особь своего вида левым глазом проявляется и в этих типах социальных взаимодействий. Играя друг с другом, детеныши одного возраста часто объединяются в пары. Мы обнаружили, что при приближении к другому детенышу сайгачата предпочитают смотреть на потенциального партнера по играм левым глазом.
Наблюдения за молодыми самцами показали, что во время «тренировочных» турниров самцы предпочитают поворачивать голову таким образом, чтобы противник находился в поле зрения левого глаза. Так же самцы располагаются в пространстве во время погони за своим противником после турнира. Эти результаты демонстрируют, что зрительная латерализация, проявляющаяся в поведении детенышей при контактах с матерью, не специфическая особенность материнско-детских взаимоотношений. Предпочтение воспринимать информацию о себе подобных левым глазом (правым полушарием) детеныши проявляют по отношению к разным социальным объектам, и такую же латерализацию демонстрируют взрослые особи. Результаты нашей работы еще раз доказывают, что асимметрия мозга и поведения не уникальна для человека, это общая черта позвоночных животных [14].
Авторы благодарят сотрудников заказника «Степной» и его директора В. Г. Калмыкова, а также Л. А. Колпащикова («Объединенная дирекция заповедников Таймыра») за помощь в организации исследования.
Исследования, проведенные на северном олене, выполнены при поддержке Российского научного фонда (проект 14-14-00284), на сайгаке — при поддержке Российского фонда фундаментальных исследований (проект 14-04-31390).
Литература
1. Бианки В. Л. Механизмы парного мозга. Л., 1989.
2. Vallortigara G., Chiandetti C., Sovrano V. A. Brain asymmetry (animal) // WIREs Cogn. Sci. 2011. V. 2. P. 146–157.
3. Бианки В. Л. Асимметрия мозга животных. Л., 1985.
4. Nottebohm F., Stikes T. M., Leonard C. M. Central control of song in the canary, Serinus canarius // J. Comp. Neurol. 1976. V. 165. P. 457–486.
5. Rogers L. J., Anson J. M. Lateralization of function in the chicken forebrain // Pharmacol. Biochem. Behav. 1979. V. 10. P. 679–686.
6. Denenberg V. H. Hemispheric laterality in animals and the effects of early experience // Behav. Brain Sci. 1981. V. 4. P. 1–49. doi: 10.1017/S0140525X00007330.
7. MacNeilage P. F. Implications of primate functional asymmetries for the evolution of cerebral hemispheric specializations // Primate laterality: current behavioral evidence of primate asymmetries / Eds J. P. Ward, W. D. Hopkins. N.Y., 1993. P. 319–341.
8. MacNeilage P. F., Rogers L. J., Vallortigara G. Origins of the left and right brain // Sci. Amer. 2009. V. 301. P. 60–67.
9. Ströckens F., Güntürkün O., Ocklenburg S. Limb preferences in non-human vertebrates // Laterality. 2013. V. 18. P. 536–575. doi: 10.1080/1357650X.2012.723008.
10. Deng C., Rogers L. J. Prehatching visual experience and lateralisation in the visual Wulst of the chick // Behav. Brain Res. 2002. V. 134. P. 375–385.
11. Rogers L. J. Lateralization in vertebrates: its early evolution, general pattern and development // Advances in the study of behavior / Eds P. J. B. Slater, J. Rosenblatt, C. Snowdon, T. Roper. San Diego, 2002. P. 107–162.
12. Rogers L. J. Early experiential effects on laterality: research on chicks has relevance to other species // Laterality. 1997. V. 2. P. 199–219. doi: 10.1080/713754277.
13. Giljov A. N., Karenina K. A., Malashichev Y. B. An eye for a worm: Lateralisation of feeding behaviour in aquatic anamniotes // Laterality. 2009. V. 14. P. 273–286. doi: 10.1080/13576500802379665.
14. Vallortigara G., Rogers L. J. Survival with an asymmetrical brain: advantages and disadvantages of cerebral lateralization // Behav. Brain Sci. 2005. V. 28. P. 575–589. doi: 10.1017/S0140525X05370105.
15. Bisazza A., Brown C. Lateralization of cognitive functions in fish // Fish cognition and behavior / Eds C. Brown, K. Laland, J. Krause. Oxford, 2011. P. 298–324.
16. Brancucci A., Lucci G., Mazzatenta A., Tommasi L. Asymmetries of the human social brain in the visual, auditory and chemical modalities // Phil. Trans. R. Soc B: Biol. Sci. 2009. V. 364. P. 895–914. doi: 10.1098/rstb.2008.0279.
17. Rosa-Salva O., Regolin L., Mascalzoni E. et al. Cerebral and behavioural asymmetries in animal social recognition // Comp. Cogn. & Behav. Rev. 2012. V. 7. P. 110–138.
18. Austin N. P., Rogers L. J. Limb preferences and lateralization of aggression, reactivity and vigilance in feral horses, Equus caballus // Anim. Behav. 2012. V. 83. P. 239–247.
19. Harris L. J. Side biases for holding and carrying infants: reports from the past and possible lessons for today // Laterality. 2010. V. 15. P. 56–135.
20. Manning J. T., Heaton R., Chamberlain A. T. Left-side cradling: similarities and differences between apes and humans // J. Human Evol. 1994. V. 26. P. 77–83.
21. Sieratzki J. S., Woll B. Neuropsychological and neuropsychiatric perspectives on maternal cradling preferences // Epidem. Psichiatr. Soc. 2002. V. 11. P. 170–176.
22. Vervloed M. P., Hendriks A. W., Eijnde E. van den. The effects of mothers’ past infant-holding preferences on their adult children’s face processing lateralisation // Brain Cogn. 2011. V. 75. P. 248–254.
23. Каренина К. А., Гилёв А. Н. Мать и дитя: Поведенческая асимметрия в природе // Наука из первых рук. 2013. Т. 5/6. С. 110–123.
24. Каренина К. А., Гилёв А. Н., Малашичев Е. Б. Асимметрия пространственного взаиморасположения матери и детеныша у сайгака (Saiga tatarica) // Материалы 3-й научной конференции «Поведение и поведенческая экология млекопитающих». Черноголовка, 14–18 апреля 2014. М., 2014. С. 47.
Межполушарная асимметрия головного мозга человека и способности детей
Введение
Как известно, экология – это наука, изучающая связи живых организмов с окружающей средой. Все связи образуют единую и очень сложную систему, которую мы называем жизнью на Земле. Человечество – неотъемлемая ее часть. А самое важное и дорогое в его жизни – здоровье, состояние которого зависит от множества факторов или их совокупности. Мозг – пожалуй, самый важный орган человека. Он управляет нашим телом, отвечает за наше сознание и эмоции. Природа, создав наш мозг как единый управляющий орган, наделила его полушария неодинаковыми способностями и обязанностями. Установление взгляда на головной мозг, как на орган психической деятельности можно причислить к величайшим научным открытиям человечества.
За последние двадцать лет изучение организации и деятельности мозга продвигалось ускоренными темпами. В прошедшее десятилетие удалось найти методы, с помощью которых можно выявлять взаимоотношения между различными частями этого органа. Интеллектуальный и творческий потенциал личности зависит от бесконечного множества факторов, особая комбинация которых и дает определенный результат. Одним из таковых факторов является функциональная асимметрия и специализация полушарий а также их интегральное взаимодействие, которое определяет не только интеллект, но и многие функциональные особенности организма. Именно рассмотрению этого явления посвящена данная работа.
Актуальность
Зачастую мы слышим от учителей: «Ты же способный, можешь иметь хорошие оценки по всем предметам», или « Ты не успеваешь на «5» по рисованию», хотя по всем предметам у ученика пятёрки. Не всегда желания учителя, родителей, учеников совпадают с возможностями самого ученика.
Обзор литературы
Мозг – это целостная морфо-функциональная система, все звенья которой одновременно, но с разными скоростями на протяжении жизни человека созревают и перекомбинируют свои внутренние связи в зависимости от доминирующих по характеру задач на том или ином возрастном периоде или в той или иной конкретной ситуации. Работа мозга, как органа, отражающего огромное число параметров внешнего мира и внутренней среды организма, в норме носит упорядоченный характер, подчиняющийся ряду законов и принципов.
Установление взгляда на головной мозг, как на орган психической деятельности можно причислить к величайшим научным открытиям человечества. За последние двадцать лет изучение организации и деятельности мозга продвигалось ускоренными темпами. В прошедшее десятилетие удалось найти методы, с помощью которых можно выявлять взаимоотношения между различными частями этого органа. Интеллектуальный и творческий потенциал личности зависит от бесконечного множества факторов, особая комбинация которых и дает определеный результат. Одним из таковых факторов является функциональная асимметрия и специализация полушарий а также их интегральное взаимодействие, которое определяет не только интеллект, но и многие функциональные особенности организма. С функциями левого и правого полушария у человека связаны два типа мышления — абстрактно-логическое и пространственно-образное
Было показано, что правое полушарие, создающее специфический пространственно-образный контекст, имеет решающее значение для творчества. Так, при органическом поражении левого полушария мозга у художников и музыкантов практически не страдают их артистические способности, а иногда даже повышается уровень эстетической выразительности творчества, но поражения правого полушария способны привести к полной утрате способности к творчеству.
Асимметрия мозга тесно связана с полом. Среди детей леворуких, заик, косоглазых, дислектиков, невротиков, страдающих недержанием мочи и кала на каждую девочку приходится около пяти мальчиков. Известно, что существует определенная зависимость между этими явлениями, и все они тесно связаны с асимметрией мозга. Например, при насильственном переучивании леворуких детей письму правой рукой у них часто появляются перечисленные аномалии, умственная отсталость, психозы, дефекты речи. Представления о половых различиях в работе мозга основаны, прежде всего, на результатах клинических и поведенческих исследований. При повреждениях левого полушария в результате кровоизлияния, опухоли или при оперативном удалении части височной доли, по поводу эпилепсии, дефицит вербальных функций у мужчин бывает гораздо больше, чем у женщин. Аналогичные повреждения правого полушария также приводят к большему дефициту функций невербального характера у мужчин, по сравнению с женщинами. Афазия вследствие н, и имеет более тяжелый характер. Поэтому был сделан вывод, что у женщин языковые и пространственные способности представленповреждения левого полушария возникает у мужчин в три раза чаще, чем у женщиы более симметрично, чем у мужчин.
Психологический половой диморфизм — разные способности и склонности мужчин и женщин, разная профессиональная пригодность и предпочтение, разная обучаемость и сообразительность — может быть связан как с половыми различиями в латерализации мозга, так и с социальными факторами. Например, по вербальным способностям: речи в целом, скорости и беглости речи, правописанию, навыкам чтения, кратковременной памяти, конформности мышления во всех возрастных группах выше уровень у женщин.У женщин гораздо лучше развито и с возрастом меньше атрофируется обоняние. У мужчин сильнее развиты пространственно-зрительные способности. Мальчики в школе значительно лучше девочек понимают геометрические концепции, эти различия меньше по алгебре, и ещё меньше по арифметике. В технических вузах максимальные преимущества мужчин также по начертательной геометрии. Они лучше ориентируются в визуальных и тактильных лабиринтах, лучше читают географические карты, легче определяют левое и правое. В шахматах, в музыкальной композиции, изобретательстве и другой творческой деятельности мужчины достигают успеха существенно чаще, чем женщины. Мала доля женщин среди сатириков, юмористов, карикатуристов, комиков и клоунов. У мужчин значительно чаще, чем у женщин, обнаруживается также преимущество правого уха при дихотическом прослушивании и левой руки у праворуких при тактильном узнавании предметов дигаптическим методом. Обнаружен половой диморфизм по соотношению длин левой и правой височных плоскостей. Половые различия отмечаются в анатомических, клинических, дихотических, тахистоскопических, электрофизиологических и психологических исследованиях полушарий.
Полный материал смотреть во вложении
Функциональная асимметрия мозга и эмоции
Значение функциональной асимметрии мозга для регуляции эмоций — проблема, привлекающая внимание многих специалистов. За последние 25-30 лет появилась огромная и труднообозримая литература на эту тему. Я не буду излагать ни историю вопроса, ни суммировать имеющиеся по этой проблеме концепции, теории, гипотезы и точки зрения. Я просто расскажу о тех материалах, которые накопились за долгие годы работы в нашей лаборатории.
Для изучения роли функциональной асимметрии мозга в организации эмоциональной сферы существует несколько путей.
- Мы можем исследовать изменения эмоционального состояния испытуемых, когда изменено состояние одного из полушарий. Это хорошо известный и многократно испытанный путь исследования больных с локальными поражениями правого и левого полушарий, при очаговой эпилепсии, это исследование изменений эмоционального состояния при пробе Вада (интракаротидном введении барбамила) или после унилатерального припадка (УП).
- Другой, прямо противоположный путь исследования — это тестирование функционального состояния каждого полушария при отчетливо выраженных эмоциональных состояниях — очень плохом (депрессивном) или патологически повышенном (маниакальном).
- Есть еще один путь — посмотреть, как изменяется деятельность . каждого полушария при направленной модификации эмоционального состояния под влиянием психотропных препаратов. Этому будет посвящена специальная лекция.
Анализ эмоциональных изменений после унилатеральных электросудорожных припадков
Как я говорил, первый путь — изучение эмоциональных изменений, когда изменено состояние одного из полушарий. Удобной моделью является анализ эмоциональных изменений после УП, которые вызывают преходящее угнетение одного полушария. Анализ проведен на больных, у которых чередовались левосторонние и правосторонние УП в течение одного курса ЭСТ. У части больных, кроме того, применялись и билатеральные электрошоки. Наиболее часто мы использовали височную унилатеральную позицию электродов (80%), реже — лобную позицию (20%).
Речь пойдет об изменениях настроения, которые становятся явными после восстановления сознания. Нас интересовало не само по себе эмоциональное состояние, а его изменение по сравнению с доприпадочным, т.е. эмоциональный сдвиг.
На рис. 3-1 на круговых диаграммах представлена частота эмоциональных сдвигов. Заметно, что эмоциональные сдвиги развиваются в половине всех случаев. При билатеральных припадках положительные и отрицательные сдвиги одинаково часты. После УП знак сдвига связан со стороной воздействия. После правосторонних УП в основном наблюдаются позитивные сдвиги, улучшение настроения. Снижение настроения происходит очень редко, в единичных случаях. После левосторонних УП возникает противоположная картина: в основном наблюдается отрицательный эмоциональный сдвиг и лишь в единичных случаях — положительный. У больных с инвертированной доминантностью связь знака эмоций и стороны воздействия отутствует. И после правосторонних, и после левосторонних УП положительные и отрицательные сдвиги встречаются одинаково часто. Частота эмоциональных сдвигов зависит от нозологии. При депрессиях они наблюдаются вдвое чаще, чем при шизофрении (60% и 30% соответственно). У больных с шизофреническим дефектом и эмоциональным обеднением эмоциональные сдвиги встречаются крайне редко (8%).
Наиболее развернутая картина эмоциональных сдвигов наблюдается после височных УП (Деглин, Николаенко, 1975). Клиника зависит от исходных состояний и синдрома. У депрессивных больных после правосторонних УП смягчается или исчезает меланхолическая симптоматика — тоска, чувство вины, суицидные мысли. Тревожные больные успокаиваются, у ипохондричных больных редуцируются неприятные ощущения. Такое улучшение отражается не только в высказываниях больных, но и в двигательной активности, позе, мимике. Сдвиг может не ограничиваться смягчением исходной симптоматики, но иногда развивается даже гипоманиакальное состояние со всеми типичными его признаками.
У больных шизофренией положительный эмоциональный сдвиг после правосторонних УП также проявляется смягчением исходной симптоматики — исчезает бредовая настороженность, подозрительность, улучшается контакт, больные перестают скрывать патологические переживания. И у больных шизофренией наблюдается чрезмерный подъем настроения, но в этом случае он часто имеет мориоподобный гебефренический характер.
Отрицательный эмоциональный сдвиг после левосторонних УП также зависит от исходной симптоматики. У депрессивных больных усиливается меланхолия, суицидные мысли становятся настойчивее, тревога нарастает вплоть до ажитации. Больные шизофренией после левостороннего УП становятся более злобными, подозрительными, нарастает негативизм. Наиболее типичным эффектом левосторонних УП является чувство общего дискомфорта.
Эмоциональные сдвиги после лобных УП сходны по клинике с височными, хотя все проявления менее выражены. Но после лобных УП наблюдается еще один эффект, который практически не наблюдается после височных УП. Это острый эмоциональный паралич, остро наступающая эмоциональная индифферентность, тупость. Этот эффект наблюдается примерно в 25% всех случаев одинаково часто после правосторонних и левосторонних УП.
Примечательной особенностью сдвигов является их динамика. Они становятся явными после прояснения сознания. Максимальной выраженности достигают постепенно и сглаживаются также постепенно. Чаще всего эмоциональные сдвиги сохраняются в течение 5-7 часов после процедуры, но нередко длятся сутки и более. Я хочу подчеркнуть, что своей длительностью эмоциональные сдвиги отличаются от всех других эффектов УП, которые сглаживаются обычно в течение 1 часа. Более того, после абортивных припадков, когда сознание, речь, моторика и другие психические функции восстанавливаются мгновенно, эмоциональные сдвиги могут развиваться столь же ярко, как после развернутых судорог и сохраняться столь же долго.
Главный факт, таким образом, заключается в том, что угнетение правого полушария сопровождается положительным эмоциональным сдвигом, а угнетение левого полушария — ухудшением настроения, отрицательным эмоциональным сдвигом.
Однако, как я говорил, эмоциональные сдвиги развиваются после УП примерно в половине случаев. Мы попытались выяснить, почему одностороннее воздействие не всегда вызывает эмоциональное изменение (Деглин, Николаенко, 1975). С этой целью у 40 больных была проведена регистрация ЭЭГ в течение всего послеприпадочного периода. В послешоковом периоде наблюдалась межполушарная асимметрия — в раздражавшемся полушарии доминирует высокоамплитудная дельта-активность (2-4 колебания в секунду), а альфа-ритм хотя и регистрируется, но редуцирован. В интактном полушарии доминирует альфа-ритм, иногда гиперсинхронный, а дельта-ритм менее выражен, чем в раздражавшемся. Мы сопоставили ЭЭГ и эмоциональные изменения после УП. Оказалось, что существует высокодостоверная статистическая связь между возникновением эмоционального сдвига после электрошока и наличием межполушарной асимметрии ЭЭГ. Иными словами, эмоциональный сдвиг развивается в тех случаях, когда вследствие УП изменяются межполушарные отношения и раздражавшееся полушарие оказывается менее активным, чем интактное. В тех случаях, когда УП не вызывает достаточно выраженного изменения межполушарных отношений, вероятность возникновения эмоциональных сдвигов невелика.
Далее мы рассмотрели, как соотносится эмоциональный сдвиг с выраженностью дельта- и альфа-ритма в каждом полушарии, и тут нас ждал сюрприз. Оказалось, что положительный эмоциональный сдвиг после правостороннего УП связан не с угнетением правого полушария, а с реципрокной активацией левого полушария. После левосторонних УП ухудшение настроения наблюдается, когда активность левого полушария снижена, а правого — повышена. Таким образом, эмоциональные сдвиги зависят в первую очередь от степени активации левого полушария.
Подведем краткий итог. При изучении эффектов право- и левосторонних УП мы столкнулись с латерализацией эмоциональных механизмов: угнетение правого полушария сопровождается улучшением настроения, угнетение левого полушария сопровождается ухудшением настроения. Анализ послеприпадочных ЭЭГ показал, что важным условием возникновения эмоциональных сдвигов служит появление межполушарной асимметрии биоэлектрической активности, то есть дисбаланс активации полушарий, причем главную роль в этом играет левое полушарие. Положительный эмоциональный сдвиг при угнетении правого полушария оказывается следствием реципрокной активации левого полушария. Отрицательный эмоциональный сдвиг при угнетении левого полушария оказывается следствием его угнетения.
Однако я высказываю эти соображения в виде гипотезы, так как, естественно, имело бы смысл повторить исследование данного вопроса на более совершенном техническом уровне и более тщательно.
Обнаруженные эмоциональные сдвиги независимы от других нарушений, возникающих в послешоковом периоде: изменения настроения развиваются и достигают максимума, когда речевые, гностические и двигательные нарушения уже отсутствуют. Более того, эмоциональные сдвиги происходят и после абортивных припадков, когда иные нарушения высших функций вообще не возникают или когда они мимолетны. Иначе говоря, изменения настроения являются самостоятельным феноменом, связанным с изменением межполушарных взаимоотношений, и не могут рассматриваться как следствие других нарушений.
Для лобных УП наиболее характерным феноменом является возникновение эмоциональной индифферентности, острый эмоциональный паралич. По-видимому, сохранность лобных структур необходима для поддержания эмоционального потенциала. Я допускаю, что лобные структуры имеют особое отношение к регуляции межполушарных отношений и дефицит их функций может привести к ригидности межполушарных взаимоотношений. Если согласиться с тем, что ригидность межполушарных отношений является важным условием регуляции эмоциональной сферы, то ригидность межполушарных взаимоотношений должна сопровождаться эмоциональным оскудением и уплощением. Еще старые авторы много раз отмечали недостаточность лобных структур в далеко зашедших случаях шизофрении. Не в этом ли причина основного проявления шизофренического дефекта — эмоциональной тупости?
Последнее, что я хочу отметить в этом разделе лекции, — это длительность эмоциональных сдвигов: до суток и более. Никакие другие послеприпадочные феномены не могут сравниться с эмоциональными по их длительности. Очевидно, одностороннее электрошоковое воздействие лишь запускает цепь реакций. Вероятно, перестройка межполушарных взаимоотношений — лишь начальный момент такой цепи реакций.
Асимметрия полушарий мозга при эмоциональной патологии
Второй, прямо противоположный путь изучения роли функциональной активности мозга в регуляции эмоций — это тестирование функционального состояния каждого полушария в случаях, когда сильно выражено то или иное эмоциональное состояние. Естественно, удобной моделью таких выраженных эмоциональных состояний являются стойкие патологические изменения настроения — депрессии и маниакальные состояния.
Наиболее сложной проблемой при таком изучении является методическая Как охарактеризовать функциональное состояние или степень активации каждого полушария? Традиционно эта проблема решалась с помощью ЭЭГ исследований. Мы отказались от электрофизиологических методов. С их помощью было проведено немало исследований, а разброс фактических данных и их интерпретаций оказался достаточно велик. В такой ситуации имело смысл искать новые, не базирующиеся на анализе биоэлектрической активности методы исследования Мы остановились на измерении некоторых параметров слуховых и зрительных функций, которые отражают степень активированности каждого полушария. Первой методикой стало измерение постстимуляционной слуховой адаптации (ПССА). Известно, что после сильной звуковой нагрузки, громкого звука наблюдается потеря слуха — преходящее повышение абсолютных слуховых порогов. В нашей лаборатории были исследованы механьзмы слуховой адаптации и было показано, что она регулируется неспецифичными активирующими влияниями стволовых структур (Балонов, Деглин, Кауфман, 1975). Чем выше активация — тем сильнее и длительнее потеря слуха после звуковой нагрузки, чем ниже активация — тем слабее и короче потеря слуха. В исследовании ПССА после УП мы показали, что такая активация раздельна для каждого полушария Моноауральное измерение слуховой адаптации позволяет судить о степени активированное™ ипсилатерального полушария. Таким образом, измерение адаптации оказалось достаточно простым способом тестирования каждого полушария в отдельности.
Измерение ПССА проводится следующим образом. Определяется абсолютный порог слуха на частоте 1000 Гц, затем на это ухо дается дозированная слуховая нагрузка — тот же тон 1000 Гц — 90 дБ над порогом в течение 1,5 минут, и сразу после окончания нагрузки несколько раз измеряется абсолютный порог для этого же тона. Разница в величине абсолютных порогов до и после нагрузки показывает величину потери слуха, а время возвращения к первоначальному порогу — ее длительность. Оба эти параметра характеризуют слуховую адаптацию, а значит и уровень активации ипсилатерального полушария.
Второй метод оценки функционального состояния полушария базируется на измерении порогов зрительных последовательных образов.
Последовательным образом (ПО) называется быстро проходящие зрительные ощущения после исчезновения стимула. Л.Я.Балонов показал, что ПО также зависят от неспецифической активации мозга: чем сильнее восходящие неспецифические влияния, тем легче возникают, ярче и длительнее сохраняются зрительные ПО (Балонов, 1972). Мой сотрудник А.Ю.Егоров исследовал «однополушарные» ПО, возникающие при стимуляции одного поля зрения. Он показал, что чем сильнее активировано полушарие, тем ниже порог возникновения ПО в ипсилатеральном этому полушарию поле зрения; чем слабее активация полушария, тем выше порог возникновения ПО (Егоров, 1986).
Измерение порогов проводится следующим образом. Тахистоско-пически в одном поле зрения предъявляется с предпороговых экспозиций стимул (черная линия на белом фоне), постепенно экспозиция увеличивается, пока после окончания стимула не появится ПО (белая линия на черном фоне). Минимальная экспозиция в миллисекундах, при которой появляется ПО, принимается за порог. Таким образом, и этот метод является достаточно простым способом количественной оценки активации каждого полушария.
Обратимся теперь к результатам, полученным при изучении активации правого и левого полушарий мозга у депрессивных и маниакальных больных. Систематическое исследование этого вопроса было проведено моей аспиранткой М.САсадовой (Асадова, 1985; 1987).
На рис. 3-2 представлены результаты измерений адаптации и последовательных образов у больных с депрессией и маниакальным состоянием по сравнению со здоровыми. Сперва обратимся к результатам, полученным на здоровых испытуемых. Видно, что межполушарной асимметрии нет — адаптация одинакова при измерениях на обоих ушах. Отсутствует также и асимметрия по порогам возникновения ПО в полях зрения. Таким образом, и ПССА, и пороги возникновения ПО свидетельствуют о том, что у здоровых испытуемых активность обоих полушарий одинакова и хорошо сбалансирована.
У больных с выраженной депрессией имеет место значительная асимметрия ПССА (рис. 3-2Б). При измерениях на правом ухе она усилена — вдвое увеличилась потеря слуха, втрое ее длительность после звуковой нагрузки. На левом ухе величина адаптации такая же, как у здоровых. Это говорит о значительном преобладании активации правого полушария. При нормализации настроения наблюдается сглаживание асимметрии. Пороги возникновения ПО также оказались асимметричными: пороги в правых полях зрения вдвое ниже, чем в левых. Это также свидетельствует о преобладании активации правого полушария. Другой факт состоит в общем повышении порогов ПО. В целом, они в несколько раз выше, чем в норме. Такое общее повышение порогов свидетельствует о значительном общем снижении активации обоих полушарий. Итак, данные измерений свидетельствуют о том, что депрессия характеризуется общим снижением активации обоих полушарий. При этом наблюдается межполушарный дисбаланс — преобладает активация правого полушария.
В отличие от депрессивных, у маниакальных больных наблюдается противоположная картина (рис. 3-2В). ПССА повышена на левом ухе, то есть преобладает активация левого полушария. Пороги возникновения ПО очень низкие в обоих полях, но в левом поле зрения они в пять раз ниже, чем у здоровых, и в несколько десятков раз ниже, чем у депрессивных больных, что свидетельствует об общей высокой активации полушарий. При этом имеет место значимая асимметрия. Пороги в левом поле зрения вдвое ниже, чем в правом. При нормализации настроения мы опять видим выравнивание асимметрии.
Суммируя результаты измерений, мы имеем основания утверждать, что изменения настроения сопровождаются общими изменениями активации полушарий и дисбалансом межполушарной активации. При депрессии наблюдается общее снижение активации полушарий, причем особенно снижена активация левого полушария. При маниакальном состоянии наблюдается общее повышение активации полушарий, причем особенно повышена активация левого полушария.
Наши старые данные, полученные при изучении УП, свидетельствуют о том, что общемозговая активация в значительной степени зависит от состояния левого полушария. При угнетении левого полушария снижается общий уровень бодрствования. Есть основания полагать, что существует особенно тесная связь между левым полушарием и активирующими структурами ствола мозга (Николаенко, 1975). С этой точки зрения, результаты данной серии исследований могут быть интерпретированы следующим образом. Знак эмоционального состояния — повышение или понижение настроения — определяется межполушарным балансом активации. Если преобладает активация правого полушария, то наблюдается плохое настроение. Если преобладает активация левого полушария, то наблюдается хорошее настроение. Однако сам межполушарный баланс активации зависит от состояния левого полушария. Преобладание активности правого полушария при депрессии является следствием снижения активации левого полушария. В то же время преобладание активности левого полушария при маниакальном состоянии связано с повышением его активации.
Таким образом, по данным исследования и депрессивных, и маниакальных состояний, можно полагать, что решающую роль в регуляции настроения играет левое полушарие. Избирательное повышение или снижение его активности определяет направление эмоциональных изменений. Ноя хочу «реабилитировать» правое полушарие и посвящу ему последнюю часть лекции.
Правое полушарие и эмоциональная экспрессия
До сих пор я говорил об эмоциях как о субъективных состояниях. Однако есть другой аспект характеристики эмоций, которого я не касался, — эмоциональная экспрессия: внешнее выражение собственных эмоций и восприятие эмоциональной экспрессии окружающих. Нужно сказать, что этой проблеме уделено много внимания в литературе, и в этом вопросе единодушия гораздо больше, чем в остальных, касающихся эмоций. Подавляющее большинство авторов отдает пальму первенства правому полушарию.
Все наши данные получены после проведения УП (Балонов, Деглин, Николаенко, 1984). Уже первые наблюдения показали различный характер изменений эмоциональной экспрессии после правосторонних и левосторонних УП. После правосторонних УП часто наблюдается гипомимия: утрачивается эмоциональная выразительность лица, оно выглядит безразличным, как бы «разглаженным». Одновременно утрачивается или снижается эмоциональная выразительность голоса — речь становится монотонной, тусклой, голос перестает отражать эмоциональное состояние, утрачиваются эмоциональные оттенки речи. После левосторонних УП нарушений эмоциональной экспрессии не наблюдается, скорее может выявиться некоторая ее утрированность — как голосовая, так и мимическая. На рис. 3-4 видно, что до УП и после левосторонних УП нарушений ни интонационной, ни лицевой выразительности нет. После правосторонних УП они наблюдаются в половине всех случаев.
Для исследования опознания эмоциональных интонаций предъявлялись магнитофонные записи 20 квази-фраз, состоящих из бессмысленных слов, но произнесенных с утрированной интонацией — гнева, радости, удивления, печали, раздражения. Больные должны были определить характер эмоции, с которой произнесена квази-фраза.
В контроле до УП приблизительно 3/4 всех ответов были правильными. В условиях угнетения правого полушария опознание зрительных интонаций резко ухудшилось. Больные напряженно вслушивались, пожимали плечами, жаловались, что «не разобрать», «непонятно» и т.д. Количество правильных ответов снизилось до половины, но в большинстве случаев ответ давался только после наводящих вопросов («Это похоже на радость?», «Это похоже на гнев?»), с колебаниями, сомнениями, большим латентным периодом. После левосторонних УП количество правильных ответов составило свыше 90% и было значимо больше, чем у этих же больных в контроле, причем ответы давались быстро, четко, уверенно.
Для исследования опознания эмоций по выражению лица был использован набор фотографий лиц с утрированной эмоциональной мимикой: радости, восторга, печали, горя, злобы, страха, ужаса, брезгливости, отвращения, высокомерия, удивления, изумления. В контрольных исследованиях до УП правильно опознавались эмоции на большинстве фотографий. После правосторонних УП опознание эмоций по выражению лица резко ухудшалось. Правильно квалифицировались меньше половины предъявленных фотографий В основном опознавались (далеко не всегда) только наиболее выраженные степени некоторых эмоциональных состояний (злобы, ужаса). Градации одних и тех же эмоций не улавливались. В некоторых случаях опознание вообще оказывалось невозможным. При предъявлении лиц даже с сильно выраженными эмоциями больные отвечали: «обыкновенное выражение», «ничего особенного» и т.д. После левосторонних УП нарушения опознания отсутствовали, правильных ответов было даже больше, чем в контроле (и причем значительно).
Таким образом, и в порождении эмоциональной экспрессии, и в ее восприятии главную роль играет правое полушарие.
В целом, наше исследование показало, что эмоция, характеризующаяся единством субъективного состояния и внешнего выражения, может расслаиваться. Субъективное эмоциональное состояние зависит от соотношения активности двух полушарий. Активация левого полушария и сопряженное снижение активности правого полушария сопровождается улучшением настроения. Снижение активности левого полушария и сопряженная активация правого полушария сопровождаются ухудшением настроения. Таким образом, по нашим данным, основную роль в регуляции эмоционального состояния играет левое полушарие, в первую очередь его височные структуры. Наши данные свидетельствуют, что лобные структуры обоих полушарий важны для сохранения эмоционального потенциала. Возможно, их функция заключается в регуляции эмоционального взаимодействия, а дефицит их функций ведет к ригидности межполушарных отношений и, как следствие, к эмоциональному обеднению. Если для субъективного эмоционального состояния решающую роль играет уровень активности левого полушария, то для эмоциональной экспрессии — уровень активности правого полушария. Угнетение правого полушария сопровождается нарушением собственной эмоциональной экспрессии (мимической и голосовой) и ухудшением опознания мимической и голосовой экспрессии окружающих. Активация правого полушария сопровождается усилением экспрессии и улучшением опознания экспрессии других людей.
В заключение я хочу сказать, что проблема связи функциональной асимметрии и эмоций очень сложна. Суждения, касающиеся этой проблемы, неоднозначны. Приведенные мною факты попадают в окружение других фактов, полученных другими авторами, и могут в силу этого трактоваться иначе. Однако я твердо уверен в одном — целостное эмоциональное поведение требует участия обоих полушарий.
Асимметрия мозга, функциональные аспекты | SpringerLink
Раздел
1
Цитаты- 342
Загрузки
Часть
Чтения из Энциклопедии нейробиологии
серия книг (REN)
Реферат
Функциональная асимметрия двух полушарий головного мозга является важной характеристикой организации и познания человеческого мозга.Это явление также называют церебральным доминированием, функциональной или латерализацией мозга и полушарной специализацией. Все эти термины относятся к тому факту, что правое и левое полушария играют разные роли в опосредовании поведения и высших психических процессов или познания. В общих чертах, речь, язык и практические навыки (двигательное планирование) зависят от функционирования левого полушария, тогда как пространственные навыки зависят от обработки правого полушария. Функциональная асимметрия существует не только у взрослых, но также у маленьких детей и младенцев и, по крайней мере, в некоторой степени, у некоторых нечеловеческих видов.
Ключевые слова
Повреждение левого полушария головного мозга Предпочтение рук Вербальный стимул Сильвиева трещина
Эти ключевые слова были добавлены машиной, а не авторами. Это экспериментальный процесс, и ключевые слова могут обновляться по мере улучшения алгоритма обучения.
Это предварительный просмотр содержимого подписки,
войдите в
, чтобы проверить доступ.
Предварительный просмотр
Невозможно отобразить предварительный просмотр. Скачать превью PDF.
Дополнительная литература
Bradshaw JL, Nettleton NC (1983):
Церебральная асимметрия человека
.Нью-Джерси: Prentice-Hall
Google Scholar
Galaburda AM и др. (1978): Человеческий мозг: цитоархитектонические лево-правые асимметрии в височной области речи.
Arch Neurol
35: 812–817
CrossRefGoogle Scholar
Witelson SF (1977): Анатомическая асимметрия височных долей: ее документация, филогенез и связь с функциональной асимметрией.
Ann NY Acad Sci
299: 328–354
CrossRefGoogle Scholar
Witelson SF (1985): О специализации полушария и пластичности мозга с рождения: Mark II In:
Функция полушария и сотрудничество у ребенка
, Best C, ed.Нью-Йорк: Academic Press
Google Scholar
Witelson SF (1985): Мозговые связи: мозолистое тело больше у левшей.
Наука
229: 665–668.
CrossRefGoogle Scholar
Информация об авторских правах
© Springer Science + Business Media New York 1988
Авторы и аффилированные лица
Нет данных о принадлежности
Асимметрия мозга — обзор
8 Симметрия и асимметрия
Асимметрия мозга на фоне фундаментальной двусторонней симметрии мозга и тела.Мы принадлежим к типу, известному как bilateria, древнему происхождению, которое включает более 1,5 миллиона современных видов животных, в том числе таких разнообразных существ, как почвенные нематоды, плодовые мухи и млекопитающие. Считается, что появление этой линии знаменует переход от неподвижных или дрейфующих планктонных животных к активным пловцам и нолам. Окаменелости билатерального периода датируются примерно 600 миллионами лет назад, задолго до кембрия (Chen et al., 2004). Двусторонняя симметрия может даже предшествовать bilateria, поскольку она также присутствует у некоторых видов филума cnidaria, который находится вне bilateria.Например, у морского анемона Nematostella vectensis двусторонняя симметрия зависит от экспрессии гомологичных генов hox так же, как и в bilateria, что позволяет предположить, что двусторонняя симметрия возникла еще до эволюционного раскола между cnidaria и bilateria (Finnerty и др., 2004). Как почти все виды, мы построены по фундаментально симметричному плану.
Однако при физической активности, которая не затрагивает напрямую сенсомоторные системы, отказ от симметрии может иметь преимущества.Асимметрия внутренних органов является наиболее очевидным прецедентом. Точно так же различные церебральные асимметрии наиболее очевидны, когда они задействованы во внутренних вычислениях, таких как язык или даже блуждание разума (Corballis, 2015), которые относительно независимы от воздействия окружающей среды. Двусторонняя симметрия кажется по сути неэффективной с вычислительной точки зрения и подразумевает дублирование функций, что является расточительным с точки зрения нейронного пространства. Эти различные преимущества асимметричной организации, тем не менее, должны быть сопоставлены с симметричными адаптациями к сенсомоторной активности.
Некоторые исследования показывают, что билатеральность может быть невыгодной по сравнению с асимметрией. Отсутствие церебрального доминирования, что отражается в смешанной манере поведения, долгое время ассоциировалось с языковыми проблемами, включая дислексию и заикание (Foundas et al., 2003; Orton, 1937), хотя доказательства часто были двусмысленными. Исследование Bishop et al. (2014) указывает, что отсутствие латерализации левого полушария не является причиной языковых нарушений у детей, но может быть его маркером. Одно крупномасштабное исследование 11-летних подростков показало снижение академической успеваемости, в том числе вербальных, новербальных, математических и читательских способностей у смешанных рук по сравнению с левшами или правшами (Crow et al., 1998). Эти авторы называют амбидекстричность «точкой полушарной нерешительности» — симметричный мозг, так сказать, не может принять решение. Однако этот вывод был поставлен под сомнение (Mayringer and Wimmer, 2002).
Исследования также предполагают, что смешанная правота связана с большей чувствительностью к сенсорным иллюзиям (Niebauer et al., 2002) и более высоким риском шизофрении (Claridge et al., 1998; DeLisi et al., 2002; Orr et al. , 1999; Upadhyay et al., 2004) и шизотипии (Somers et al., 2009; Tsuang et al., 2013), а также другие проблемы психического здоровья (Rodriguez, Waldenstrom, 2008; Rodriguez et al., 2010).
Твердая вера в паранормальные явления, по-видимому, связана с симметричной мозговой активностью (Pizzagalli et al., 2000). Люди, получившие высокие баллы по магическому мышлению, практически не различаются между полями зрения в задаче лексического решения, что подразумевает отсутствие церебрального доминирования, в то время как те, у кого низкий балл, демонстрируют преимущество левого поля зрения, тогда как те, кто набирает относительно высокие баллы, не показывают разницы между полями зрения (Pizzagalli et al., 2001). По сравнению с левшами и правшами, смешанные руки также демонстрируют более высокую склонность к магическим идеям (Бадзакова-Трайков и др., 2011; Barnett and Corballis, 2002). Этот эффект, хотя, кажется, применим к ручности, определяемой с точки зрения предпочтения, и не был очевиден в исследовании, в котором ручность определялась с точки зрения относительной производительности при выполнении задачи по касанию пальцем (Grimshaw et al., 2008).
Принимая во внимание этот перечень очевидных недостатков, какие компенсирующие преимущества могут быть получены в результате двусторонних отношений? Визуализация мозга предполагает, что креативность больше зависит от сотрудничества между полушариями, чем от одного полушария (Lindell, 2011), и есть некоторые свидетельства того, что смешанные руки могут демонстрировать более высокий уровень креативности, чем правши или левши (Бадзакова-Трайков и другие., 2011; Shobe et al., 2009). Многие выдающиеся спортсмены и спортсмены обладают смешанным доминирующим положением и могут извлечь пользу из дополнительной степени перцептивного и моторного баланса. Аннетт (2002) также утверждает, что среди хирургов непропорционально большое количество несправедливостей. Однако сама леворукость часто ассоциируется с отсутствием церебральной асимметрии, а не с ее обращением вспять, и очень много исключительных людей были левшами, в том числе пять недавних президентов США (Форд, Рейган, Джордж Х.У. Буш, Клинтон, Обама).
Два человека, которые могут служить символами беспристрастного, — это Леонардо да Винчи и Альберт Эйнштейн. Леонардо — прототип «человека эпохи Возрождения», художник, ученый и изобретатель — обычно считается человеком, который на несколько веков раньше своего времени. Он был левшой и обычно писал задом наперед, в зеркальном письме, но он также был способен писать нормально. Эйнштейн, похоже, был правшой, но, как говорят, он медленно развивал речь и учился медленно, а посмертный анализ его мозга выявил «необычную симметрию между полушариями» (Witelson et al., 1999, стр. 2151; см. также Falk, 2009; Falk et al., 2013), особенно в затылочной и теменной долях. Сообщается также, что он сказал: «Я хочу знать, как Бог создал этот мир».
Некоторые из предполагаемых недостатков, связанных с отсутствием церебрального доминирования или отклонениями от праворукости, вполне могут свидетельствовать об исключительных качествах, способных к адаптации способами, не очевидными в стандартных тестах или традиционном понимании. Это поднимает вопрос о том, действительно ли расстройства, связанные с латерализацией, такие как паранормальные переживания, галлюцинации и магические представления, являются «расстройствами» или просто частью нормальной сети человеческого существования.Даже психические заболевания могут быть адаптивными, или когда-то так было. Кауфман (2016) указывает, что когда-то галлюцинации считались нормальным явлением и играли роль в жизни провидцев, таких как Иисус из Назарета, Святой Павел из Тарса и даже Сократ, и предполагает, что это произошло благодаря сочинениям Вольтера. , Дарвин и Фрейд, что они стали ассоциироваться с психическими заболеваниями. Творчество тоже давно ассоциируется с шизофренией и биполярными расстройствами, и исследования также предполагают генетическую связь (Power et al., 2015).
Джейнс (1976) предположил, что церебральная асимметрия возникла во втором тысячелетии до нашей эры в ответ на различные катастрофы, такие как наводнения, вторжения и тому подобное. До этого людьми управляли галлюцинации, взывающие к богам, но церебральная асимметрия позволяла левому полушарию создавать ощущение себя, так что люди брали на себя ответственность за свои действия. Теория Джейнса имеет мало эволюционного смысла, поскольку, как предполагалось ранее, ручность и церебральная асимметрия почти наверняка предшествовали возникновению нашего вида.Тем не менее, в идее о том, что церебральная асимметрия лежит в основе рационального мышления, вполне может быть доля правды, и что отсутствие асимметрии вполне может привести к более бредовым и, возможно, галлюцинаторным мыслительным процессам, которые все еще частично поддерживают религиозное мышление. Компромисс между симметрией и асимметрией может иметь отношение к вековой борьбе между религией и наукой, примером которой является религиозное противостояние таким светилам науки, как Чарльз Дарвин и Галилей.
Магическое мышление само по себе может быть связано с творчеством и иметь положительные последствия для науки и математики.Леонард и Брюггер (1988) отмечают связь между паранормальными мыслями, бредовыми мыслями и творчеством и предполагают, что эти характеристики связаны с повышенной активацией правого полушария и относительно грубой семантической активацией в этом полушарии. Это, в свою очередь, приводит к ослаблению ассоциаций и повышению творческих способностей. Хотя работа Леонарда и Браггера сосредоточена на правом полушарии, возможно, что этот профиль связан с отсутствием церебрального доминирования, а не с какой-либо специализацией самого правого полушария.Несмотря на доказательства Crow et al. (1998), что у смешанных рук не хватает математических способностей, Сингх и О’Бойл (2004) сообщают, что математически одаренные подростки не демонстрируют асимметрии полушария при выполнении задач, связанных с глобально-локальными суждениями и сопоставлением букв, тогда как подростки со средними способностями и студенты колледжей демонстрируют преимущество левого полушария, предполагающее, что у математически одаренных людей может отсутствовать последовательная церебральная асимметрия. Хотя Сингх и О’Бойл выбрали для этого исследования правшей, они также характеризуют математически одаренных как «типично мужчин, левшей и близоруких» (стр.371).
Этот умозрительный ход мысли, кажется, возвращает нас к двойному мозгу, за исключением того, что дуальность возникает между симметрией и асимметрией, а не между левым и правым полушариями. Учитывая многомерную природу асимметрии мозга и сложность генетических влияний, различия, несомненно, все еще более сложны, чем можно уловить с помощью дихотомии или даже одного измерения. Более общий момент заключается в том, что латеральность является источником вариаций, которые сами по себе являются адаптивными.Вариация — это сама эволюция, и даже внутри нашего собственного вида вариации в поведении, познании и личности обеспечивают эффективную социальную жизнь, позволяя людям выполнять множество специализированных ролей. Сатмари (2015) пишет, что язык, сам по себе сильно латерализованный и подверженный индивидуальным изменениям, был одним из семи основных переходов в эволюции, предлагая нечто беспрецедентное — «согласованное разделение труда». Но это разделение, столь важное для эволюции сложных обществ, зависит не только от языка, но и от индивидуальных различий в других областях, включая пространственные способности, креативность, атлетизм и вычислительные способности.
Асимметрия головного мозга — обзор
4.62.3.3.2 Воздействие неонатальной новизны и мозг
Влияние воздействия неонатальной новизны на мозг изучалось на уровне функциональной асимметрии мозга, общей анатомии гиппокампа и синаптической пластичности. Давно известно, что ранняя стимуляция, такая как процедура манипуляции, вызывает асимметричные изменения в мозге (Denenberg, 1981). В частности, у необработанных крыс повреждение левой и правой коры головного мозга оказывает схожее влияние на активность открытого поля, в то время как крысы, которых лечили, демонстрируют дифференциальные изменения в ответ на повреждение левой и правой коры (Denenberg, 1978).Учитывая этот асимметричный эффект, исследование влияния неонатальной новизны на развитие мозга характеризовалось отдельными измерениями для левого и правого полушарий, тогда как большинство исследований ранней стимуляции не различали левое и правое измерения.
В соответствии с этим открытием асимметричного эффекта ранней стимуляции, новые и домашние крысы различаются по своей маневренности, причем новые крысы демонстрируют сдвиг влево в предпочтении лапы при выполнении задачи (Tang and Verstynen, 2002).Этот сдвиг влево постоянно наблюдается в течение последовательных дней и в течение нескольких месяцев задержки, что свидетельствует о стойком усилении правого церебрального доминирования. Эта интерпретация подтверждается еще одним надежным наблюдением второй формы измерения функциональной асимметрии мозга, поворотной асимметрией во время спонтанного исследования в новой среде. При первом входе в новую среду новые крысы имеют больший уклон вправо, чем домашние крысы (Tang et al., 2003b). Поскольку поворот вправо включает более сильное толчок левой передней конечности, эта разница в асимметрии также указывает на усиление правого церебрального доминирования (Tang and Reeb, 2004).В отличие от функциональной асимметрии руки, эта асимметрия поворота временна и, по-видимому, зависит от новизны ситуации. Например, он показывает явную модуляцию во времени, причем наибольший эффект новизны обнаруживается в первые минуты спонтанного исследования (Tang et al., Неопубликованное наблюдение). В совокупности эти результаты представляют собой четкие доказательства модификации асимметрии мозга в результате воздействия неонатальной новизны.
Поскольку гиппокамп играет решающую роль в обучении и памяти (Eichenbaum, 1997), а также в регулировании выработки HPA (Herman et al., 2003; Sapolsky et al., 1984a), были исследованы эффекты воздействия неонатальной новизны на анатомию и функцию этой структуры. Хотя нет общих различий в объеме гиппокампа, новые и домашние крысы различаются по своим моделям объемной асимметрии, причем новые крысы демонстрируют относительно больший правый гиппокамп, чем домашние крысы, на 1% от общего объема гиппокампа (Verstynen et al., 2001). Хотя 1% кажется небольшим эффектом, с вычислительной точки зрения он может существенно повлиять на динамику нейронных сетей, вызывая функциональную асимметрию (Reggia et al., 1998).
Влияние воздействия новизны на синаптическую пластичность было исследовано в области СА1 гиппокампа. Долгосрочная потенциация (ДП), наиболее изученная форма синаптической пластичности, различается у новых и домашних крыс. Независимо от стороны гиппокампа, воздействие неонатальной новизны приводит к усилению LTP, в то время как синаптическая передача до индукции LTP не показывает значительной разницы (Tang and Zou, 2002). При дальнейшем анализе данных с учетом стороны гиппокампа обнаруживаются доказательства асимметричных эффектов воздействия новизны.Этот эффект стимуляции в раннем возрасте характеризуется двумя формами селективности: селективностью в отношении правого гиппокампа и селективностью в отношении LTP (Tang et al., 2008).
Поскольку все эти асимметричные изменения в мозге связаны с усилением доминирования функции правой стороны, возникает вопрос, как достигаются эти асимметричные эффекты и какое эволюционное значение может иметь эта асимметричная среда. Поскольку показатели асимметрии, как было показано, обладают прогностической силой для показателей памяти (Tang et al., 2003a; Tang and Reeb, 2004) и асимметрия синаптической пластичности избирательна для LTP, можно предположить, что ранний опыт критически важен для развития нормальной функциональной латерализации или предотвращения аномальной функциональной латерализации. Следует отметить, что аномальная функциональная латерализация связана с рядом психопатологий (Davidson, 2003).
Полушарная асимметрия анатомической сети человеческого мозга, выявленная диффузионно-тензорной трактографией
Топологическая архитектура церебральной анатомической сети отражает структурную организацию человеческого мозга.Недавно топологические меры, основанные на теории графов, предоставили новые подходы для количественной оценки крупномасштабных анатомических сетей. Однако мало исследований изучали асимметрию полушарий человеческого мозга с точки зрения сетевой модели, и мало что известно об асимметрии паттернов соединений областей мозга, которые могут отражать функциональную интеграцию и взаимодействие между различными областями. Здесь мы использовали визуализацию тензора диффузии для построения бинарных анатомических сетей для 72 здоровых взрослых субъектов-правшей.Мы установили наличие структурных связей между любой парой из 90 корковых и подкорковых областей с помощью детерминированной трактографии. Чтобы исследовать асимметрию полушарий мозга, был проведен статистический анализ, чтобы выявить области мозга со значительными различиями между двусторонними топологическими свойствами, такими как степень связности, характерная длина пути и центральность промежуточности. Кроме того, были исследованы локальные структурные связи для изучения локальной асимметрии некоторых специфических трактов белого вещества.С точки зрения как глобальных, так и локальных паттернов связи мы идентифицировали области мозга с полушарной асимметрией. В сочетании с предыдущими исследованиями мы предположили, что топологическая асимметрия в анатомической сети может отражать функциональную латерализацию человеческого мозга.
1. Введение
Мозг демонстрирует асимметрию как в макроскопической структуре, так и в микроскопической цитоархитектуре. Более того, многие исследования выявили анатомическую асимметрию, соответствующую функциональной латерализации [1–3].Например, левосторонняя асимметрия в размере областей участвует в речевой и слуховой обработке, таких как височная плоскость [4–7], сильвиевая щель [8] и извилина Хешля [4, 9, 10], что соответствует левополушарному каналу. доминирование языка [11]. Недавно функциональные нейровизуализационные исследования установили специфичность полушария для ряда языковых, моторных и пространственных задач [12]. Структурные МРТ-исследования выявили структурную асимметрию в толщине коры или объемах серого вещества различных областей мозга [13, 14].Используя методы диффузионной тензорной визуализации (DTI), исследователи также идентифицировали левостороннюю асимметрию некоторых трактов белого вещества, таких как пучки цингулюма [15, 16] и дугообразный пучок [2, 3, 17–21].
Эти предыдущие исследования изучали структурную или функциональную асимметрию некоторых конкретных областей мозга или анатомические связи между ними. Асимметрия серого или белого вещества была проанализирована по местным региональным признакам. Недавно сетевая модель была предложена как полезный инструмент для исследования структурной организации и функциональных механизмов человеческого мозга [22–31].Теория графов подходит к анализу сложных сетей, которые могут предоставить новый мощный способ количественной оценки структурных и функциональных систем мозга. С помощью сетевых моделей все больше и больше исследований показывают, что структурные и функциональные сети человеческого мозга обладают атрибутами маленького мира [23–26, 28, 30, 32–34] и модульной структурой [35–37]. В нескольких исследованиях с использованием диффузионной МРТ были предложены различные методы построения анатомической сети мозга [25, 32, 33, 36, 38, 39].Все эти исследования показали, что корковые сети человеческого мозга имеют топологию «маленького мира», которая характеризуется большими коэффициентами кластеризации и короткой средней длиной пути [30, 40]. Однако мало исследований изучали асимметрию полушарий с точки зрения церебральной анатомической сети, и мало что известно об асимметрии паттернов соединений областей мозга, которые могут отражать функциональную интеграцию и взаимодействие между различными областями.
В этом исследовании мы сначала построили анатомическую сеть для каждого субъекта с помощью метода детерминированной диффузной тензорной трактографии (DTT), а затем применили подходы теории графов для изучения топологических свойств двусторонних областей мозга в сети.Чтобы исследовать асимметрию полушарий мозга, был проведен статистический анализ, чтобы выявить области мозга со значительными различиями между двусторонними топологическими свойствами. Кроме того, были исследованы локальные структурные связи для изучения локальной асимметрии некоторых специфических трактов белого вещества. С точки зрения как глобальных, так и локальных паттернов связи мы идентифицировали области мозга с полушарной асимметрией.
2. Материалы и методы
2.1. Субъекты
В это исследование были включены 72 здоровых взрослых субъекта (42 мужчины; средний возраст; средний год обучения). Все участники были правши согласно Эдинбургской инвентаризации [41]. Каждый участник предоставил письменное информированное согласие перед обследованием МРТ, и это исследование было одобрено Комитетом по этике медицинских исследований больницы Сюаньу Столичного медицинского университета.
2.2. Сбор данных
DTI был выполнен с помощью системы 3T Siemens Trio MR с использованием стандартной головной катушки.Движение головы было минимизировано с помощью удерживающих подушек из поролона, предоставленных производителем. Диффузионно-взвешенные изображения были получены с использованием последовательности однократной эхо-планарной визуализации (EPI), совмещенной с передне-задней комиссуральной плоскостью. Методика интегрального параллельного сбора данных (iPAT) использовалась с коэффициентом ускорения 2. Время сбора данных и искажение изображения из-за артефактов восприимчивости можно уменьшить с помощью метода iPAT. Градиенты, повышающие чувствительность к диффузии, применялись вдоль 12 нелинейных направлений (= 1000 с / мм 2 ) вместе с получением без взвешивания диффузии (= 0 с / мм 2 ).Параметры визуализации: 45 непрерывных аксиальных срезов с толщиной среза 3 мм и без зазора, поле зрения = 256 мм × 256 мм, время повторения / время эхо = 6000/87 мс и матрица сбора данных = 128 × 128. Реконструкция матрица была 256 × 256, что давало разрешение в плоскости 1 мм × 1 мм. Для каждого участника также было получено сагиттальное T1-взвешенное трехмерное изображение с использованием подготовленной намагниченностью последовательности быстрого градиентного эхо-сигнала (MP-RAGE). Параметрами визуализации для этого были поле зрения 22 см, время повторения / время эхо = 24/6 мс, угол поворота = 35 ° и размеры вокселя 1 мм × 1 мм × 1 мм.
2.3. Предварительная обработка данных
Вихретоковые искажения и артефакты движения в наборе данных DTI были скорректированы путем применения аффинного выравнивания каждого взвешенного по диффузии изображения к изображению = 0 с использованием FMRIB’s Diffusion Toolbox (FSL, версия 3.3; http: //www.fmrib. ox.ac.uk/fsl). После этого процесса, элементы тензора диффузии были оценены путем решения уравнения Стейскала и Таннера [42, 43], а затем восстановленная матрица тензора была диагонализована для получения трех собственных значений (,, и) и собственных векторов.Дробная анизотропия (FA) каждого вокселя рассчитывалась по следующей формуле: DTT был реализован с помощью DTIstudio, версия 2.40, программное обеспечение (http://www.mristudio.org), с использованием метода «назначение волокна путем непрерывного отслеживания». [44]. Все участки в наборе данных были рассчитаны путем заполнения каждого вокселя с FA более 0,2. Трактография прекращалась, если она поворачивалась на угол более 50 градусов или достигала воксела с FA менее 0,2 [45].
2.4. Построение анатомической сети
Мы построили анатомическую сеть для каждого пациента на основе оптоволоконной связи из детерминированного DTT.Основные процедуры заключаются в следующем. Во-первых, мозг был автоматически сегментирован на 90 корковых и подкорковых областей (по 45 для каждого полушария; см. Таблицу 1) с помощью шаблона AAL (подробности см. [32]). Вкратце, отдельные T1-взвешенные изображения были сопоставлены с изображениями b0 в пространстве DTI. Затем преобразованные изображения T1 были нелинейно преобразованы в шаблон ICBM152 T1 в пространстве Монреальского неврологического института (MNI). Обратные преобразования использовались для деформации атласа AAL из пространства MNI в собственное пространство DTI.Используя эту процедуру, мы получили 90 корковых и подкорковых областей интереса, каждая из которых представляет собой узел сети. Во-вторых, все волокна в головном мозге были получены с помощью детерминированной волоконной трактографии. И тогда два узла и соединяются ребром, если существует не менее трех волокон с концами в областях и; порог трех волокон был выбран, чтобы гарантировать, что средний размер самого большого подключенного компонента сети составляет 90 для всех субъектов. Количество волокон между областями использовалось только для обозначения наличия / отсутствия края.Таким образом, бинаризованная анатомическая сеть для каждого субъекта была построена и представлена симметричной матрицей 90 × 90.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2.5. Сетевой анализ
Мы исследовали топологические свойства анатомической сети на региональных (узловых) уровнях. Региональные свойства были описаны с точки зрения степени (), длины кратчайшего пути () и промежуточной центральности () узла. Здесь мы даем краткие формальные определения каждого узлового свойства, использованного в этом исследовании.
2.5.1. Степень
Степень узла определяется как количество подключений к этому узлу. Узлы с высокой степенью связи имеют большую степень.Степень графа — это среднее значение степеней всех узлов в графе: это мера для оценки степени разреженности сети.
2.5.2. Длина кратчайшего пути
Средняя длина кратчайшего пути узла — это наименьшее количество ребер, которые необходимо пройти, чтобы установить соединение между узлом и узлом. Характерная длина пути сети — это среднее значение длины кратчайшего пути между узлами: количественно оценивает возможность параллельного распространения информации или глобальную эффективность (в терминах) сети [47].
2.5.3. Централизованность посредничества
Центральность посредничества широко используется для идентификации наиболее центральных узлов в сети, которые связаны с теми узлами, которые действуют как мосты между другими узлами. Расстояние между узлами определяется как количество кратчайших путей между парами других узлов, которые проходят через узел [48, 49]. Затем рассчитывалась нормализованная промежуточность: Узлы с наибольшими нормализованными значениями промежуточности считались ключевыми узлами (т.е., хабы) в сети.
2.6. Реконструкция трактов белого вещества
Для дальнейшего исследования локальной асимметрии структурных связей мы затем реконструировали несколько основных трактов белого вещества, соединяющих различные области мозга. Основываясь на анатомических знаниях о проекциях волокон, в нескольких исследованиях предложены протоколы отслеживания основных трактов белого вещества [21, 50, 51]. Согласно опубликованным протоколам отслеживания, мы реконструировали двусторонние пучки поясной извилины (CB), оптическое излучение (OR), нижний лобно-затылочный пучок (IFO), нижний продольный пучок (ILF), дугообразный пучок (AF) и крючковидный пучок (UF) для каждого из них. предмет.На основе реконструированных трактов для каждого субъекта была рассчитана средняя FA каждого волоконного тракта путем усреднения значений FA по вокселям, которые образуют трехмерные тракты, полученные из трактографии.
2.7. Анализ асимметрии
Чтобы проанализировать различия полушарий в топологических свойствах областей мозга, мы вычислили коэффициент латеральности для каждого свойства (, и). Мы проверили недействительность этого отношения в группе, используя непараметрический односторонний знаковый критерий (после поправки Бонферрони для множественных сравнений; i.е., 90/2 = 45 пар регионов). Чтобы проанализировать различия полушарий в структурных свойствах трактов белого вещества, мы сравнили средние значения FA и количество волокон в каждом тракте между левым и правым полушариями с помощью парных -тестов. Для каждого тракта была определена значительная асимметрия FA или числа волокон, если. Все статистические анализы были выполнены с помощью Matlab.
3. Результаты
3.1. Области мозга с полушарной асимметрией в свойствах узлов
На основе бинарной анатомической сети, построенной для каждого субъекта, мы вычислили топологические свойства (, и) каждого узла для каждого субъекта.Путем статистического анализа для всех субъектов мы выявили некоторые области с асимметрией полушарий в свойствах узлов (рис. 1). Мы определили левую асимметрию с лучшими топологическими свойствами в левом полушарии, чем в правом, например, больше и меньше в левом полушарии. Точно так же правая асимметрия определялась как области с большим и меньшим размером в правом полушарии. По результатам мы выявили некоторые области с асимметрией полушарий по всем трем топологическим свойствам, такие как левосторонняя асимметрия в нижней лобно-треугольной извилине, островке, нижней теменной извилине и задней медиальной коре (парацентральная долька, предклинье и задняя поясная извилина) и вправо. асимметрии верхней лобной извилины, гиппокампа, верхней теменной извилины, надмаргинальной извилины, угловой извилины и среднего височного полюса (после коррекции Бонферрони).
3.2. Структурная асимметрия трактов белого вещества
Для каждого субъекта мы можем успешно реконструировать большинство двусторонних трактов белого вещества (рис. 2 (а)). Однако для некоторых объектов трудно отследить правый автофокус (15 из 72). Из рисунка 2 (b) мы можем видеть, что несколько участков белого вещества демонстрируют полусферическую асимметрию как в микроструктурных (значение FA), так и в макроструктурных (число волокон) свойствах, таких как CB, ILF и AF (, нескорректировано).
4.Обсуждение
В этом исследовании мы исследовали асимметрию полушарий человеческого мозга с точки зрения церебральной анатомической сети, построенной на основе данных DTI. Сравнивая двусторонние топологические свойства, мы выявили некоторые области мозга со значительной асимметрией влево или вправо, что указывало на асимметричные паттерны соединения этих областей. Более того, структурные свойства некоторых локальных трактов белого вещества также обнаруживают полушарную асимметрию, что указывает на асимметрию локальных связей.Он предположил, что структурные организации человеческого мозга асимметричны как по отношению к глобальным, так и локальным паттернам связей. Затем следует обсудить функциональное значение этих структурных асимметрий.
4.1. Полушарные асимметрии в узловых свойствах анатомической сети
Предыдущие исследования полушарных асимметрий в человеческом мозге были сосредоточены на структурах или функциях некоторых локальных областей [1, 2, 20, 52, 53]. В этом исследовании мы исследовали асимметрию полушарий на основе паттернов связи между различными областями мозга, сравнивая топологические свойства узлов между двусторонними полушариями в анатомической сети.
Различные узловые свойства отражают разные аспекты узла в сети. В этом исследовании мы выбрали три топологических свойства, степень, нормализованную центральность между узлами и длину кратчайшего пути, чтобы проанализировать полушарные асимметрии анатомической сети. Степень означает количество прямых подключений к узлу. Большая степень означает более структурные связи с другими областями мозга в бинарной анатомической сети. Центральность между посредничеством отражает важность узла, и узел с высокой центральностью, таким образом, имеет решающее значение для эффективной коммуникации [48, 49].Самая короткая длина пути количественно определяет параллельное распространение информации или глобальную эффективность (в терминах) узла, а меньшая означает более высокую глобальную эффективность параллельной передачи информации [47]. Хотя эти три свойства взаимосвязаны друг с другом, они отражают разные аспекты узла в сети. Следовательно, узел с большей степенью, более высокой центральностью и меньшей длиной кратчайшего пути будет играть более важную роль в сети. Затем мы предположили, что асимметричные свойства областей в двусторонних полушариях указывают на латерализацию этих областей в анатомической сети.
Поскольку были выявлены области с полушарной асимметрией в узловых свойствах, мы классифицировали эти области по их функциям следующим образом.
Области с левосторонней асимметрией (1) Язык и слуховая функция: средняя и нижняя височная извилина [54, 55], хвостатое ядро [56, 57], извилина Хешля [58, 59], а также треугольная и глазничная часть нижней лобная извилина [60, 61]. (2) Зрительная функция: средняя и нижняя височная извилина [62, 63], калькариновая щель и окружающая кора [62, 64].(3) Эмоции, ощущения и зависимость: островок [65, 66]. (4) Ассоциативная кора: парацентральная долька, предклинье, задняя поясная извилина и нижняя теменная извилина.
Области с правой асимметрией (1) Пространственное внимание: угловая и надмаргинальная извилина [67–69]. (2) Распознавание лица: веретенообразная извилина [70, 71]. (3) Эмоции и память: гиппокамп и миндалевидное тело [68 , 72–74]. (4) Ассоциация коры: верхняя и средняя лобная извилины, верхняя теменная извилина и средний височный полюс.
Из результатов видно, что области с левосторонней асимметрией в основном связаны с языком, визуальной обработкой и сенсорными функциями.Области с асимметрией вправо в основном связаны с функциями пространственного внимания, распознавания лиц, эмоций и памяти. Некоторые области ассоциативной коры с множественными функциями также демонстрируют левую или правую асимметрию узловых свойств.
В сочетании с результатами некоторых предыдущих исследований мы предположили, что топологическая асимметрия в анатомической сети, вероятно, формирует структурный субстрат для различных функциональных принципов обработки информации в двух полушариях.С момента открытия Пола Брока в 1861 году понятие языковой специализации левого полушария утвердилось [11]. В последнее время все больше и больше исследований выявляют левостороннюю асимметрию в размерах областей, участвующих в речевой и слуховой обработке, таких как планум височная [4–7], сильвиевая щель [8] и извилина Хешля [4, 9, 10], которые может служить анатомической основой функциональной латерализации человеческого мозга. В этом исследовании наиболее очевидной находкой является левосторонняя асимметрия в нескольких областях, связанных с языком, таких как треугольные и орбитальные области нижней лобной извилины, средней и нижней височной извилины и извилины Хешля.Результаты совпадают с преобладающим представлением о том, что левое полушарие является доминирующим полушарием для речи [75, 76]. Это также может указывать на то, что не только структурная асимметрия этих регионов, но также асимметричные паттерны соединений этих регионов поддерживают функциональную латерализацию человеческого мозга.
Другой находкой является асимметрия вправо угловой и надмаргинальной извилины, которая расположена в височно-теменном соединении. Прямоугольная и надмаргинальная извилины широко участвуют в функциях пространственного внимания [67–69].Предыдущие функциональные МРТ-исследования также показали, что правое височно-теменное соединение играет доминирующую роль в фактической реализации пространственного внимания с помощью функционального анализа связности [77, 78]. Следовательно, этот результат соответствовал специализации правого полушария для зрительно-пространственных функций [75, 79–81].
Некоторые подкорковые структуры в лимбической системе, такие как гиппокамп и миндалевидное тело, также были обнаружены с правосторонней асимметрией топологических свойств. Гиппокамп играет важную роль в памяти и пространственной навигации [68, 74], а миндалевидное тело играет главную роль в обработке памяти и эмоций [72, 73].Затем, в этом исследовании, правосторонняя асимметрия топологических свойств гиппокампа и миндалины может указывать на то, что правое полушарие более заметно в функциях эмоций и памяти. Это также согласуется с результатами некоторых структурных МРТ-исследований, которые показали, что гиппокамп и миндалевидное тело имеют правую асимметрию на основе измерений объема [82].
Помимо приведенных выше результатов, мы выявили некоторые области с полушарной асимметрией по всем трем топологическим свойствам, такие как левосторонняя асимметрия в трех областях задней медиальной коры (парацентральная долька, предклинье и задняя поясная извилина), нижней теменной извилине, островке и островке. правые асимметрии среднего височного полюса, верхней теменной извилины и верхней лобной извилины.Большинство из этих регионов расположены в ассоциативной коре головного мозга, которая играет центральную роль в получении конвергентных входных сигналов от множества корковых регионов [68]. Следует отметить, что с помощью диффузионного МРТ-исследования были выявлены три непрерывных области в задней медиальной части коры головного мозга как структурное ядро коры головного мозга [36]. Однако ни одно исследование еще не изучило функциональную или структурную асимметрию этих основных регионов. Затем в этом исследовании впервые выявляется левосторонняя асимметрия основных регионов анатомической сети.
Следует отметить, что аномальные асимметричные паттерны структуры или функции мозга связаны с некоторыми психическими расстройствами, такими как шизофрения, а степень измененной асимметрии связана с симптомами пациентов [83]. Поэтому мы предположили, что асимметричная топология мозговых сетей также будет меняться в различных условиях с психическими заболеваниями и может служить в качестве чувствительных биомаркеров для раннего обнаружения заболеваний, что требует дальнейшего изучения.
4.2. Структурная асимметрия трактов белого вещества
Основываясь на результатах трактографии шести основных трактов белого вещества, мы проанализировали структурные асимметрии этих трактов в средних значениях ФА и количестве волокон. Предыдущие исследования DTI выявили анатомическую асимметрию некоторых трактов волокон, такую как левосторонняя асимметрия дугообразного пучка [2, 3, 19–21]. Как один из наиболее важных языковых путей, дугообразный пучок начинается от области Брока в нижней лобной извилине и выступает в среднюю и нижнюю височные извилины [84].В этом исследовании мы выявили левостороннюю асимметрию AF как по микро-, так и по макроструктурным свойствам. Левосторонняя асимметрия AF соответствует левосторонней асимметрии треугольной области нижней лобной извилины, средней и нижней височной извилины, выявленной при топологическом анализе анатомической сети. Эти результаты предполагают, что связанные с языком области демонстрируют левостороннюю асимметрию как от глобальных, так и от локальных паттернов анатомических связей. Мы предположили, что эти структурные асимметрии могут обеспечить анатомический субстрат языковой функциональной латерализации человеческого мозга.Помимо левой асимметрии AF, CB и ILF также асимметричны влево как по средним значениям FA, так и по количеству волокон. Пучки цингулюма были исследованы в нескольких предыдущих исследованиях DTI, и левосторонняя асимметрия целостности волокон была идентифицирована различными методами [15, 16, 85]. Нижний продольный пучок, соединяющий височную и затылочную доли, играет важную роль в зрительной памяти [86, 87] и рассматривается как косвенный путь языковой семантической обработки [88].Это первый случай, когда левосторонняя асимметрия ILF обнаружена, и это может предоставить новую информацию для будущих исследований. Нижний лобно-затылочный пучок, соединяющий задние затылочные области и орбитофронтальную область, является прямым путем языковой семантической обработки [88]. Предыдущее исследование DTI сообщило о левосторонней асимметрии целостности волокон IFO и предположило, что структурная асимметрия тракта соответствует преобладанию языка в полушарии [21].Кроме того, мы обнаружили, что оптическое излучение и крючковидный пучок асимметричны вправо по количеству волокон. Однако функциональное значение этих асимметрий следует изучить в будущих исследованиях.
Поскольку в настоящем исследовании изучались как глобальные (структурный коннектом), так и локальные (FA) показатели, некоторые результаты могут быть перекрестно проверены с помощью различных показателей. Например, мы обнаружили, что и области, связанные с языком, и тракты WM демонстрируют значительную левостороннюю асимметрию.Однако глобальные и локальные показатели могут иметь разные физиологические значения. Локальный показатель, такой как FA, отражает целостность белого вещества или согласованность ориентации волокон на микроструктурном уровне, в то время как узловые свойства коннектома мозга, такие как узловая эффективность, являются интегрированным показателем пропускной способности глобального информационного потока и связаны со всеми из них. узловые связи, которые состоят из определенного тракта или нескольких трактов вместе. Следовательно, результаты сетевого анализа могут предоставить более полную информацию, чем традиционные региональные и местные исследования на системном уровне.
4.3. Методологические вопросы
Самыми важными элементами сети являются узлы и ребра. Определение узлов и ребер имеет большое влияние на построенную сеть и результаты анализа. Поэтому нам необходимо решить некоторые методологические вопросы о том, как мы проводили строительство сети.
Сначала мы применили шаблон AAL, чтобы определить узлы для сети каждого субъекта. Шаблон AAL был взят из головного мозга одного пациента MNI [46]. Самым большим ограничением этого шаблона является отсутствие анатомической латерализации некоторых областей, например, левосторонняя асимметрия височного плана у подавляющего большинства правшей [46].Это ограничение повлияет на результаты топологической асимметрии анатомической сети в этом исследовании. В будущих исследованиях для исследования асимметрии сети мозга, чтобы исследовать асимметрию сети мозга, следует использовать более точное представление о парцелляции, которое определяется на уровне вокселов, а не на региональном уровне для разделения коры головного мозга на тысячи областей [36]. для более точной локализации асимметричных топологических организаций.
Во-вторых, мы использовали детерминированный DTT для определения краев анатомической сети.Однако проблема «пересечения волокон» является ограничением детерминированных алгоритмов трактографии, потому что отслеживание всегда останавливается, когда достигает областей пересечения волокон с низкими значениями фракционной анизотропии [89]. Это приведет к потере некоторых существующих волокон и, следовательно, некоторых краев сети. Еще одно ограничение детерминированной трактографии, особенно для пучков волокон на большие расстояния, — это ошибочные результаты отслеживания из-за шума и ограничений разрешения [89]. Для решения этой проблемы несколько исследователей использовали вероятностные алгоритмы отслеживания волокна [90–92].Моделируя распределение вероятностей ориентации волокон в вокселе, эти статистические методы могут идентифицировать оптоволоконные соединения, пропущенные методами детерминированного отслеживания. Однако количество направлений градиента в нашем наборе данных диффузии недостаточно для точной оценки функции плотности вероятности ориентации волокон. Следовательно, будущие исследования с использованием более совершенных методов диффузионной визуализации или методов трактографии могут дать более полную и точную анатомическую сеть для каждого субъекта.
Другой вопрос, связанный с выбором двоичной или взвешенной сети, требует решения. Для взвешенной сети проблема состоит в том, чтобы выбрать наиболее репрезентативный показатель структурной связности. Несколько возможных мер, таких как количество волокон, средняя длина волокна, плотность волокна и средняя доля анизотропии, могут быть выбраны в качестве меры связности [25, 38, 39, 93]. Но физиологический смысл этих мер не ясен. Также трудно проверить, какая мера наиболее точно описывает передачу информации нейронных сигналов.В этой работе мы построили бинарную сеть, просто принимая во внимание наличие / отсутствие региональных связей. Однако взвешенная сеть с правильной мерой связности может лучше отражать топологическую асимметрию сети.
Помимо вышеуказанных ограничений методологии, в будущем необходимо изучить некоторые другие важные вопросы. Во-первых, поскольку было высказано предположение о влиянии пола на топологическую организацию сетей мозга [94], в будущем следует изучить половые различия в асимметрии сети.Во-вторых, из-за относительно небольшого размера выборки в настоящем исследовании для проверки текущих результатов следует использовать другие независимые наборы данных с высококачественной МРТ и более крупными выборками, такие как наборы данных Human Connectome Project (HCP) [95].
5. Заключение
В этом исследовании мы проанализировали асимметрии полушария с точки зрения анатомической сети всего мозга и выявили топологические асимметрии некоторых областей мозга, которые указали на асимметричные паттерны соединений этих областей на глобальном уровне. .Более того, мы обнаружили структурную асимметрию некоторых локальных анатомических связей между регионами, а структурные асимметрии трактов белого вещества взаимосвязаны с топологической асимметрией областей мозга. Мы предположили, что асимметричные паттерны соединений областей мозга могут отражать функциональную латерализацию человеческого мозга.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении публикации данной статьи.
Благодарности
Эта работа была поддержана Программой 973 (грант № 2013CB837300, Ни Шу), Национальным фондом естественных наук Китая (гранты № 81471732, Ни Шу; 81101038 и 30930029, Яоу Лю), ECTRIMS- Стипендия MAGNMIS от ECTRIMS (Yaou Liu), Пекинского фонда естественных наук (грант № 7133244, Yaou Liu), программы Beijing Nova (грант № xx2013045, Yaou Liu), Пекинской группы по новым медицинским дисциплинам (грант № 100270569). , Ни Шу) и Фонды фундаментальных исследований для центральных университетов (грант №2013YB28, Ни Шу).
Произошла ошибка при настройке вашего пользовательского файла cookie
Произошла ошибка при настройке вашего пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера на прием файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie. - Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Полушарная асимметрия в слуховой обработке модуляций речевых огибающих у детей до чтения
Введение
Речевые сигналы содержат многочисленные слуховые подсказки, которые способствуют восприятию речи, но огибающая речи важна и с некоторой ограниченной спектральной информацией, почти достаточной для точной разборчивости речи (Drullman et al., 1994; Шеннон и др., 1995). Огибающая речи характеризуется амплитудными модуляциями (AM). Модуляции <50 Гц играют важную роль в идентификации и сегментации важнейших речевых единиц, таких как слоги (AM около 4 Гц) и фонемы (AM около 20 Гц) (Rosen, 1992). Теории о восприятии речи предполагают, что эти диапазоны модуляции обрабатываются асимметрично в слуховой коре, с предпочтением правого полушария для обработки модуляции скорости слога и двусторонним или левым полушарием для обработки модуляции скорости фонемы (Poeppel, 2003; Boemio et al., 2005). Более того, эта функциональная полушарная асимметрия, по-видимому, уже присутствует в младенчестве (Telkemeyer et al., 2009, 2011).
Недавно было высказано предположение, что нарушение этой функциональной асимметрии полушария может быть связано с языковыми расстройствами, такими как дислексия (Goswami, 2011). Дислексия развития — это неврологическое наследственное заболевание, которое характеризуется серьезными и постоянными проблемами чтения и правописания, которые, как полагают, вызваны трудностями при отображении фонологической структуры слов на уровне слогов и / или фонем и манипулировании ими (Vellutino et al., 2004). Недавние исследования показали, что отклонения в нейронном представлении сигналов речевой оболочки связаны с этими фонологическими проблемами. Hämäläinen et al. (2012) продемонстрировали, что по сравнению с асимметрией правого полушария у нормальных читателей, люди с дислексией демонстрируют более симметричную нейронную организацию во время обработки скорости слога. Кроме того, обнаружено, что модуляция скорости фонем менее латерализована (Lehongre et al., 2011) или снижена (Poelmans et al., 2012a) в левом полушарии у людей с дислексией.С другой стороны, некоторые исследования показали отклонения слуховой обработки у людей с дислексией уже на уровне ствола мозга (Hornickel et al., 2009).
Однако большинство этих исследований проводилось на взрослых (Lehongre et al., 2011; Hämäläinen et al., 2012; Poelmans et al., 2012a) и у детей школьного возраста с дислексией (Abrams et al., 2009), и Мало что известно о том, как и когда эти нервные изменения проявляются у детей дошкольного возраста, до того, как они научатся читать. Учитывая, что нейронная обработка модуляции скорости слога и фонемы сильно связана с фонологической осведомленностью (Abrams et al., 2009; Lehongre et al., 2011; Poelmans et al., 2012a) и восприятие речи (Alaerts et al., 2009; Poelmans et al., 2012a), возможно, измененные модели реакции на эти модуляции у предварительно читающих детей могут предсказать проблемы с чтением до того, как они возникнут.
Чтобы исследовать это, были измерены устойчивые слуховые ответы (ASSR) на модуляцию скорости слога и фонемы у детей с предварительным чтением с высоким наследственным риском дислексии (на основе родственников первой степени родства с официальным диагнозом дислексии) и у детей контрольной группы, предварительно читающих.Мы стремились изучить асимметрию полушарийной реакции на модуляции скорости слога и фонемы коры головного мозга и определить, отличаются ли дети дошкольного возраста с высоким наследственным риском дислексии от модуляции скорости слога и / или скорости фонемы по сравнению с детьми контрольной группы. Кроме того, мы стремились изучить обработку на этих корковых уровнях, а также обработку на уровне ствола мозга.
Результаты
Целью этого исследования было изучить корковое представление медленных темпоральных модуляций в речи и изучить нейронную асимметрию по отношению к модуляциям скорости слога и фонемы у детей, предварительно читающих, с высоким и низким наследственным риском дислексии.С этой целью были исследованы представления огибающей нейронной речи с помощью значений отношения сигнал / шум в ответах, а асимметрия временной обработки была исследована с помощью индексов латеральности.
Топографические карты плотности источника тока, амплитуды напряжения и формы сигналов во временной области для общего среднего значения ASSR по всем объектам для каждого условия представлены на рисунке 2.
Рис. 2. Топографические контурные карты
и формы сигналов во временной области для общего среднего значения ASSR по всем объектам для 4, 20 и 80 Гц.Карты плотности источника тока (вверху) и карты амплитудного напряжения (посередине) построены с безопорной интерполяцией и представляют собой максимальный положительный пик общего среднего усредненного периода отклика. Положение электродов обозначено красными точками. Осциллограммы во временной области (внизу) нанесены на график с эталоном Cz и представляют реакцию на четыре цикла стимула в канале P10.
Обработка нейронных слогов и фонем
Для усредненного SNR отклика по девяти электродам в каждом полушарии был проведен дисперсионный анализ ANOVA с повторными измерениями с группой (HR против LR) в качестве межсубъектного фактора и полушарием (левое полушарие против правого полушария) в качестве внутрисубъектного фактора. три тестируемых частоты модуляции по отдельности (рис.3).
Рисунок 3.
Среднее SNR ответа для детей LR (темно-серый) и HR (светло-серый) для 4, 20 и 80 Гц. Столбцы представляют собой среднее отношение сигнал / шум для девяти электродов левого и правого полушарий. Планки погрешностей показывают среднее значение ± 1 SE. *** p ≤ 0,001.
Для 4 Гц нет основного эффекта группы ( F (1,68) = 0,39, p = 0,536) или эффекта взаимодействия между группой и полушарием ( F (1,68) = 0,14 , п = 0.707), что свидетельствует об отсутствии групповых различий в отношении SNR ответа для скорости слога. В целом был обнаружен главный эффект полушария ( F (1,68) = 17,41, p <0,001). Ипсилатеральные ответы правого полушария были выше, чем ответы контралатерального левого полушария (средняя разница, 1,5 дБ; SE, 0,4 дБ). Этот основной эффект полушария был также обнаружен для отдельных электродов с помощью теста Манна – Уитни U . Для семи из девяти пар электродов ответы были значительно выше для электродов правого полушария, чем для электродов левого полушария (все p <0.037).
Для 20 Гц, нет основного эффекта группы ( F (1,69) = 0,23, p = 0,631) или полушария ( F (1,69) = 0,01, p = 0,911), ни взаимодействия ( F (1,69) = 2,52, p = 0,117) между этими факторами не наблюдалось.
Для 80 Гц нет основного эффекта группы ( F (1,70) <0,01, p = 0,946) или эффекта взаимодействия между группой и полушарием ( F (1,70) = 3 .03, p = 0,086), что свидетельствует об отсутствии групповых различий в отношении SNR ответа для ответа 80 Гц. В целом, основной эффект полушария ( F (1,70) = 218,51, p <0,001) показал, что ипсилатеральные правые ответы были значительно выше, чем контрлатеральные левые ответы (средняя разница, 4,6 дБ; SE, 0,3 дБ. ). Этот основной эффект полушария был воспроизведен на отдельных электродах с помощью теста Манна – Уитни U . Для всех пар электродов ответы правого полушария были значительно выше, чем ответы левого полушария (для всех пар электродов p <0.001).
Асимметрии временной обработки
Для дальнейшего изучения наблюдаемых основных эффектов полушария были рассчитаны LI. LI оценивались отдельно для каждой скорости модуляции и для каждой группы участников на основе теста t для одной выборки для усредненной группы электродов (рис. 4) и на основе знакового рангового теста Вилкоксона для отдельных пар электродов. Для изучения возможных различий между HR и группой LR был проведен независимый тест образца t на усредненной группе электродов, а тест Манна-Уитни U был проведен на отдельных парах электродов.
Рис. 4. Средние значения LI
для детей LR (темно-серый) и HR (светло-серый) для 4, 20 и 80 Гц. Планки погрешностей представляют собой 95% доверительный интервал среднего. ( * ) p = 0,052, * p ≤ 0,050, *** p ≤ 0,001.
Чтобы исследовать латеральность на более индивидуальном уровне, участники были разделены на категории в соответствии с их левым (LI <0) или правым (LI> 0) асимметричным ASSR на основе LI усредненной группы электродов (Таблица 2).Наконец, был проведен биномиальный тест, чтобы проверить, продемонстрировала ли значительная часть участников левую или правую асимметрию.
Таблица 2.
Участников в каждой группе (HR или LR), демонстрирующих левую или правую асимметрию ASSR для 4, 20 и 80 Гц
Для 4 Гц была обнаружена ипсилатеральная правополушарная асимметрия для группы LR (LI = 0,12, t (38) = 2,40, p = 0,022), и тенденция к правополушарной асимметрии была обнаружена для Группа HR (LI = 0.12, т (30) = 2,40, p = 0,052). Эта правополушарная асимметрия также присутствовала для большинства отдельных пар электродов (группа LR: пять из девяти пар электродов, все p <0,050; группа HR: восемь из девяти пар электродов, все p <0,050) и для > 70% людей в каждой группе участников (группа LR: 72%, z = 2,72, p = 0,001; группа HR: 71%, z = 2,34, p <0.001). Не было обнаружено существенной разницы между группами LR и HR в степени правополушарной асимметрии ( t (68) = -0,01, p = 0,995; тест U : для восьми из девяти пар электродов, p > 0,050).
Для 20 Гц ответы были симметричными для группы LR (LI = 0,04, t (38) = 1,30, p = 0,203), а также для группы HR (LI = -0,04, t (31) = -0,89, p = 0.380). Аналогичным образом симметричные ответы были обнаружены для отдельных пар электродов (группа LR: семь из девяти пар электродов, все p > 0,050; группа HR: девять из девяти пар электродов, все p > 0,050). Для 20 Гц количество участников с левой асимметрией равнялось количеству участников с правой асимметрией в группе LR (L = 44%, z = 0,80, p = 0,522), а также в группе HR (L = 53%, z = 0,35, p = 0,860). Не было обнаружено значимых групповых различий для 20 Гц ( t (69) = 1.51, p = 0,135; U тест, все p > 0,050).
Наконец, значительная ипсилатеральная правая асимметрия была обнаружена для 80 Гц для обеих групп участников для усредненной группы электродов (группа LR: LI = 0,31, t (38) = 10,98, p <0,001; группа HR : LI = 0,21, t (32) = 6,17, p <0,001), а также для отдельных пар электродов (группа LR: все p <0,001; группа HR: все p <0.001). Правая асимметрия наблюдалась у 97% детей LR ( z = 5,93, p <0,001) и 94% детей HR ( z = 5,05, p <0,001). Значительная разница в степени асимметрии была обнаружена между группами LR и HR на усредненной группе электродов ( t (70) = 2,16, p = 0,034). Однако эта разница не может быть воспроизведена на уровне отдельных пар электродов (тест U , все p > 0.050).
Обсуждение
В текущем исследовании корковые вызванные потенциалы для модуляции скорости слога и фонемы были измерены в группе детей, читающих до чтения, с наследственным риском дислексии и без нее. Дети, умеющие преподавать, демонстрировали предпочтение правого полушария при обработке модуляции скорости слога. Напротив, наблюдалась симметричная обработка модуляции скорости фонем. Никаких четких различий между группами HR и LR не наблюдалось.
В литературе приводятся доказательства функционального предпочтения полушарий скорости модуляции процесса, присутствующей во временной оболочке речи (Poeppel, 2003).Однако до сих пор нет единого мнения о точном проявлении этой асимметрии полушарийной обработки. В то время как большинство исследований показывают, что модуляция скорости слога обрабатывается преимущественно в правом полушарии (Boemio et al., 2005; Abrams et al., 2008; Hämäläinen et al., 2012), все еще ведутся споры о том, являются ли модуляции скорости звука фонем обрабатывается в левом полушарии (Abrams et al., 2008) или двусторонне (Boemio et al., 2005; Hämäläinen et al., 2012). Важным фактором, способствующим этим противоречивым результатам, является методологическая разница между исследованиями.Во-первых, в исследованиях оценивалась асимметрия с помощью различных методов записи (фМРТ: например, Boemio et al., 2005; MEG: например, Hämäläinen et al., 2012; ЭЭГ: например, Poelmans et al., 2012a). Во-вторых, факторы стимула, такие как сложность стимула (лингвистическая: например, Abrams et al., 2009; акустическая: например, Lehongre et al., 2011) или манера предъявления стимула (односторонняя или двусторонняя презентация стимула: например, Poelmans et al., 2012b ) могли повлиять на асимметрию полушария. В предыдущем исследовании мы изучали нервную чувствительность к модуляции скорости слога и фонемы у взрослых с дислексией и без нее (Poelmans et al., 2012a, b) с точно такими же стимулами и очень похожей техникой записи, которые использовались в настоящем исследовании. Мы обнаружили, что при стимуляции правого уха модуляция скорости слога и фонемы латерализовалась в правое полушарие (Poelmans et al., 2012b), и что взрослые с дислексией демонстрируют реакции нижнего левого полушария на модуляцию скорости фонемы (Poelmans et al., 2012a). ). На основании этих результатов была выдвинута гипотеза, что проблемы с грамотностью при дислексии могут быть вызваны конкретной проблемой при обработке модуляции скорости фонем в речевой оболочке.
В отличие от результатов, полученных у взрослых (Poelmans et al., 2012a), настоящее исследование не обнаружило доказательств различий в скорости обработки слогов и фонем между дошкольниками с высоким наследственным риском дислексии и детьми контрольной группы. Однако это не совсем неожиданно, учитывая неоднородность образцов, исследованных в этом исследовании. То есть текущее исследование отбирало участников на основе их наследственного риска дислексии. Тот факт, что «только» до 50% детей с высоким наследственным риском дислексии в конечном итоге разовьются дислексией (Gilger et al., 1991) подразумевает, что эта группа высокого риска очень разнородна и что последствия могут быть замаскированы. Тем не менее, в предыдущих исследованиях фМРТ структурные и функциональные изменения в задних отделах мозга были продемонстрированы у детей с высоким наследственным риском дислексии до начала чтения (Raschle et al., 2011, 2012). Для определения реальных предшественников дислексии потребуются длительное наблюдение и ретроспективное исследование текущей выборки.
В отличие от модуляции скорости слога и фонемы, для модуляции частоты управления 80 Гц было обнаружено значительное различие в абсолютной силе латерализации.Поскольку известно, что ответы 80 Гц более выражены в стволе мозга на ипсилатеральной стороне стимуляции (см. Ниже), ожидалось латеральное воздействие. Однако ответы на 80 Гц были значительно менее латерализованы вправо по ЧСС по сравнению с детьми LR. Менее латерализованный ответ у детей с ЧСС может указывать на снижение нейронной синхронизации со стимулом. Альтернативно, возможно, что меньше нейронов ствола мозга синхронно срабатывают в группе HR по сравнению с группой LR. Однако последние две возможности маловероятны, поскольку не было обнаружено различий в абсолютном SNR ответа между HR и LR детьми.Более того, наблюдаемое различие между группами HR и LR присутствовало только для усредненной группы электродов и не могло быть воспроизведено, когда анализы проводились на отдельных парах электродов. Это говорит о том, что групповая разница на 80 Гц может иметь довольно небольшой эффект. Тем не менее, в литературе нарушения слуховой обработки у людей с дислексией обнаруживаются на нескольких уровнях системы обработки слуховой информации, начиная с уровня слухового ствола мозга (например,g., Hornickel et al., 2009) до уровня слуховой коры (например, Poelmans et al., 2012a). Более того, Abrams et al. (2006) обнаружили, что точность временного кодирования речи в слуховом стволе мозга предсказывает корковую асимметрию звуков речи. Плохая нейронная синхронизация в стволе мозга может, таким образом, влиять на развитие паттернов полушарной асимметрии при модуляции скорости слога и / или фонемы.
Несмотря на отсутствие четких групповых различий, были обнаружены различные паттерны полушарной асимметрии для контрольной частоты 80 Гц, частоты слогов и модуляции частоты фонем.Для контрольной частоты 80 Гц в группах HR и LR был обнаружен правосторонний боковой ответ. Доминантные нейронные генераторы ответов 80 Гц расположены в стволе мозга до анатомического пересечения слухового пути в верхнем оливковом комплексе (Picton et al., 2003), что приводит к ипсилатеральной правой латерализации к монофонической стимуляции правого уха. Ипсилатеральная асимметрия ответов 80 Гц наблюдалась другими исследователями в исследованиях взрослых, а также в исследованиях детей и младенцев (Herdman et al., 2002; Picton et al., 2003).
Что касается скорости слога, ответы были выше в правом полушарии, чем в левом, как для групп HR, так и для групп LR. Правая асимметрия модуляции скорости слога была ранее обнаружена у взрослых (Poelmans et al., 2012a, b), детей (Abrams et al., 2009) и новорожденных (Telkemeyer et al., 2009, 2011). Вместе с настоящими открытиями это говорит о том, что специализация правого полушария для обработки модуляции скорости слога присутствует уже в очень молодом возрасте, возможно, даже с рождения.Это, в сочетании с отсутствием групповых различий в настоящем исследовании и среди взрослых (Poelmans et al., 2012a), предполагает, что маловероятно, что проблемы с чтением при дислексии вызваны нейронным отклонением в обработке модуляции скорости слога.
Что касается частоты фонем, ответы были одинаково высокими в обоих полушариях, как для групп HR, так и для групп LR. Это контрастирует с результатами, полученными у взрослых, где было обнаружено ипсилатеральное предпочтение (т. Е. Правополушарная асимметрия стимуляции правого уха) для обработки модуляции скорости фонем (Poelmans et al., 2012б). Различия между обеими возрастными группами могут иметь как структурные, так и функциональные причины. Во-первых, разница между детьми и взрослыми может быть связана с разницей в степени коркового складывания. Если активирован расширенный диапазон пирамидных клеток, ответы могут генерироваться с переменной полярностью из-за высокой складчатости, которая появляется в коре головного мозга человека (Ahlfors et al., 2010). В зависимости от локальной геометрии коры это может привести к отмене генерируемых ответов, особенно когда участки активности распространяются на противоположные стенки бороздок и извилин (Shaw et al., 2013). Более того, было показано, что степень кортикальной складчатости изменяется с возрастом и становится стабильной только в возрасте 6-7 лет (Nie et al., 2013). Следовательно, разница в степени коркового складывания между полушариями у детей и взрослых в регионе, где генерируются ответы на модуляции скорости фонем, может вызывать разные паттерны ответа полушария с этой скоростью. Во-вторых, возможно, что паттерн латерализации взрослых и детей, которые не читают, различается из-за изменений в развитии языковой сети.Подобно настоящим результатам, Telkemeyer et al. (2009, 2011) нашли доказательства симметричной обработки информации о скорости фонем у новорожденных. Учитывая, что церебральная латерализация изменяется с возрастом (Shaw et al., 2009; Bishop, 2013) и что на это развитие влияет созревание, а также опыт (Minagawa-Kawai et al., 2011), кажется, что симметричная обработка скорости фонем модуляции в предварительном чтении детей перерастают в специализированное предпочтение ипсилатерального полушария в возрасте после 5 лет.Эта увеличивающаяся кортикальная специализация для обработки модуляции скорости фонем может быть связана с фонологическим развитием и развитием чтения. Если механизмы обработки скорости фонем еще не созрели в возрасте 5 лет, это может объяснить, почему дети, которые предварительно читают, обрабатывают фонологическую информацию на уровне слога, а не спонтанно развивают понимание фонем до начала обучения чтению (Anthony et al., 2003 ; Зиглер и Госвами, 2005). Может оказаться, что для осуществления этого перехода и развития точного чтения необходима определенная степень корковой специализации.В качестве альтернативы, возможно, что явные инструкции по чтению и развитие понимания фонем имеют нисходящее влияние на развитие полушарной специализации для обработки модуляции скорости фонем в речи. Ретроспективный анализ текущего набора данных, когда выявляются дети с дислексией, может раскрыть причинную природу этих гипотез.
В этом исследовании изучали корковые вызванные потенциалы для модуляции скорости слога и фонемы в группе детей, предварительно читающих чтение, с наследственным риском дислексии и без него.Несмотря на то, что мы не обнаружили групповых различий для обработки модуляции скорости слога и фонемы, настоящие результаты предполагают незрелую модель полушарной специализации для обработки модуляции скорости фонемы у 5-летних детей. Таким образом, предполагается, что возможное нарушение в развитии полушарной специализации для обработки модуляции скорости фонем может частично лежать в основе проблем с чтением людей с дислексией. Длительное наблюдение за этими детьми проясняет это предположение.
Дифференциальный вклад двух полушарий головного мозга человека в синхронизацию действий
Функциональная асимметрия между двумя полушариями — интригующий принцип организации мозга. На поведенческом уровне они наиболее очевидны в том, как люди используют свои руки. В задачах, требующих движений обеих рук, правши обычно используют правую руку для более быстрых, динамичных движений, в то время как левая рука используется для более медленных движений или даже статического контроля положения руки (Sainburg, 2002; Swinnen and Wenderoth, 2004; Серриен и Совиярви-Спапи, 2015).Резать хлеб или забивать гвоздь в стене — повседневные примеры такой функциональной асимметрии. В лаборатории постукивание пальцем можно использовать для обнаружения асимметрии полушария, связанной с этим явлением. Как правило, правая рука отстукивает относительно более высокие частоты постукивания более точно, чем левая рука, даже при бимануальном одночастотном постукивании пальцем (Repp, 2005; Ivry, 1996; Peters, 1980). И наоборот, левая рука точнее отстукивает относительные более низкие частоты отстукивания, чем правая (Pflug et al., 2017). Это предполагает, что левое полушарие предпочтительно контролирует относительные более высокие частоты постукивания, а правое полушарие предпочтительно контролирует относительные более низкие частоты постукивания, но происхождение такой полушарной асимметрии неизвестно. Существует несколько предложений о происхождении функциональных различий между полушариями, начиная от специализированной обработки в сенсорной области, латерализованных сенсомоторных взаимодействий, асимметричного моторного контроля и до общих рамок домена полушарного доминирования (Kimura, 1993; Minagawa-Kawai et al., 2007; Тога и Томпсон, 2003; Келл и Келлер, 2016). Поведение может выиграть от параллельной обработки различных аспектов сложных стимулов и / или планирования движений в левом и правом полушарии (Serrien et al., 2006). Влиятельные теории предлагают дифференциальную сенсорную обработку относительных частот либо в спектральной, либо во временной области (Ivry and Robertson, 1998; Flevaris and Robertson, 2016; Poeppel, 2003) в качестве вычислительной основы полушарной специализации. Однако эмпирические исследования, в которых параметризовались спектральные или временные аспекты сенсорной информации, не всегда подтверждали эти теории (Луо и др., 2007; Жиро и Труи, 2002; Boemio et al., 2005). Это могло быть следствием того факта, что активность мозга лишь слегка латерализована во время задач восприятия. Однако считается, что функциональная латерализация усиливается, когда требуется моторная мощность (Ivry and Robertson, 1998; Keller and Kell, 2016).
Чтобы отделить конкретный вклад сенсорной и моторной систем в функциональную латерализацию ручного управления, мы провели два исследования изображений с использованием функциональной магнитно-резонансной томографии (фМРТ) и магнитоэнцефалографии (МЭГ).Дизайн исследования исключил, что эффекты состояния были результатом сенсорных стимулов или дифференциального использования эффекторов. В парадигме постукивания пальцами со слуховым шагом участников просили бимануально постучать по слуховым ударам. Постукивание по каждому звуковому сокращению (2,5 Гц) было определено как условие быстрого постукивания, в то время как постукивание по каждому четвертому звуковому сокращению (положение удара четыре) представляло собой медленное постукивание с частотой 0,625 Гц (см. Рисунок 1). Обе частоты попадают в естественный диапазон движений пальцев, но представляют разные концы этого спектра (Parncutt and Cohen, 1995; London, 2012; Drake and Palmer, 2000; Repp, 2003).В то время как в условиях быстрого постукивания быстрый слуховой ритм был единственным ритмом, который обрабатывался и использовался для слухомоторной синхронизации, этот более быстрый ритм служил синхронизирующим сигналом для генерации более медленного ритма в состоянии медленного постукивания. Условие медленного постукивания представляло основной интерес в нашем исследовании, потому что во время медленного постукивания два взаимосвязанных ритма должны были быть представлены параллельно, что могло потенциально выявить полушарную специализацию для управления ритмами с разными относительными частотами (Ivry and Robertson, 1998).Предыдущее поведенческое исследование (Pflug et al., 2017) показало, что представление относительно медленного ритма параллельно более быстрому должно выявить вклад правого полушария в управление руками. В то время как мы использовали фМРТ, чтобы определить, имеют ли слуховые или моторные области более выраженный профиль латерализации, который отвечает на вопрос о разном вкладе сенсорных и моторных систем в полушарную специализацию, мы использовали МЭГ для определения полушарных различий в ритмах мозга, связанных с постукиванием пальцами в метод с временным разрешением и для исследования направленной связи с временным разрешением между слуховой и моторной ассоциациями коры коры (рис. 2).
Условия врезки.
Участники были проинструктированы нажимать либо на каждый удар (быстрое нажатие, левая панель), либо на четвертую позицию из четырех последовательных слуховых ударов (медленное состояние, правая панель). Закрашенные квадраты представляют события постукивания, белые квадраты представляют слуховые удары без постукивания в состоянии медленного постукивания.
https://doi.org/10.7554/eLife.48404.003
Обзор применяемых шагов анализа МЭГ.
(1) Когерентные источники с ЭМГ-сигналом были обнаружены на высокой частоте с помощью формирователя луча DICS. (2) Временные ряды были извлечены из локализованных источников с помощью формирователя луча LCMV.(3) Был применен частотно-временной анализ со скользящим окном для преобразования этих сигналов в частотно-временное представление. Путем усреднения по частотам (14–20 Гц) был извлечен сигнал с низким бета-диапазоном. (4) Исходные сигналы подавались на анализ направленной связи с временным и частотным разрешением (TPDC).
https://doi.org/10.7554/eLife.48404.004
Известно, что движение подавляет бета-колебания и увеличивает активность в гамма-диапазоне (Muthuraman et al., 2012; Тамаш и др., 2018; Пфурчеллер и др., 2003; Пфурчеллер и Лопеш да Силва, 1999; Энгель и Фрис, 2010). Тем не менее, нервные колебания, особенно в бета-диапазоне, не только отражают текущее моторное состояние, но также участвуют во внутреннем времени, особенно во время обработки ритма, и модулируются по амплитуде во время восприятия и производства ритма не только в моторной и дополнительной моторной коре головного мозга. но также в слуховой и слуховой ассоциации коры головного мозга (Arnal and Giraud, 2012; Doelling and Poeppel, 2015; Nobre et al., 2007; Fujioka et al., 2015; Meijer et al., 2016; Morillon et al., 2014; Килавик и др., 2013; Кулашекхар и др., 2016; Морийон и Байе, 2017; Иверсен и др., 2009). Сравнение нервных колебаний во время медленного и быстрого ритмичного постукивания пальцами может показать, как мозг представляет два разных ритма параллельно. Амплитудные модуляции бета-колебаний должны различаться между функциональными гомологами в случае, если были различия в обработке полушарий во времени относительных частот постукивания.Мы предположили, что моторная и / или слуховая кора могут различаться не только по общей бета-мощности, но и по степени их представления медленных и быстрых ритмов во временной модуляции бета-мощности (Fujioka et al., 2015; Morillon and Baillet, 2017). Если предсказания гипотез, основанных на сигналах, о специализации полушария (Ivry and Robertson, 1998) верны, мы специально ожидали, что правая слуховая кора более точно будет представлять медленный ритм, а левая слуховая кора — быстрый ритм при медленном постукивании пальцами состояние, включающее оба ритма.Вместо этого доминирование левой руки в моторном контроле, основанное на навыках левостороннего секвенирования (Kimura, 1993; Haaland et al., 2004), должно было бы предсказывать контроль над обоими ритмами со стороны левого полушария. Функциональная специализация двух полушарий связана не только с латерализованной региональной активацией, но также с образованием латерализованных функциональных сетей регионов (Stephan et al., 2003; Keller and Kell, 2016). Таким образом, мы исследовали, действительно ли слухомоторные взаимодействия между правой и левой ассоциативной слуховой корой и дополнительной моторной областью (SMA), областью моторных ассоциаций, активно участвующей во внутренней генерации последовательностей (Kotz et al., 2009; Merchant et al., 2013; Merchant et al., 2015; Crowe et al., 2014), модулировались по-разному в двух полушариях, представляя медленный в дополнение к быстрому ритму. Мы предположили, что слухомоторная эффективная связь между двумя полушариями может различаться с точки зрения силы связи в бета-диапазоне. Наши результаты идентифицируют левую слуховую ассоциативную кору как первичную область коры, которая представляет относительный быстрый слуховой ритм, в то время как правая слуховая ассоциативная кора задействована для представления относительной медленной скорости постукивания в амплитудной модуляции низких бета-колебаний.Напротив, моторная кора и мозжечок представляют только временные закономерности моторной активности. Представление медленного в дополнение к быстрому ритму увеличивает функциональную связь с низким бета от правой слуховой ассоциации коры головного мозга к SMA параллельно с усилением ЖИВОЙ активации этих областей. Кроме того, более сильная и двунаправленная функциональная связь с низким бета между SMA и левой слуховой ассоциативной корой может отдавать предпочтение левому полушарию для иерархической интеграции взаимосвязанных ритмов в гештальт (Iversen et al., 2008; Суиннен и Вендерот, 2004).
.