Понятие о возбудимости. Изменения возбудимости при возбуждении. Параметры возбудимости
Возбудимостью называется способность нервной или мышечной клетки отвечать на раздражение генерацией ПД. Основным мерилом возбудимости обычно служит реобаза. Чем она ниже, тем выше возбудимость, и наоборот. Связано это с тем, что, как мы уже говорили ранее, главным условием возникновения возбуждения является достижение МП критического уровня деполяризации (Ео <= Ек). Поэтому мерилом возбудимости является разница между этими величинами (Ео — Ек). Чем меньше эта разница, тем меньшую силу надо приложить к клетке, чтобы сдвинуть мембранный потенциал до критического уровня, и, следовательно, тем больше возбудимость клетки.
Еще Пфлюгер показал, что возбудимость — величина непостоянная. Катод повышает возбудимость, анод ее понижает. Напомним, что эти изменения возбудимости под электродами носят название электротонических. Русский ученый Вериго показал, что при длительном действии постоянного тока на ткань, или при действии сильных раздражителей эти электротонические изменения возбудимости извращаются — под катодом начальное повышение возбудимости сменяется ее понижением (развивается т.н. катодическая депрессия), а под анодом сниженная возбудимость постепенно возрастает. Причина этих изменений возбудимости на полюсах постоянного тока связана с тем, что величина Ек меняется при длительном действии раздражителя. Под катодом (и при возбуждении) Ек постепенно удаляется от МП, снижается, так что наступает момент, когда разница Е0-Ек становиться больше исходной. Это приводит к падению возбудимости ткани. Напротив, под анодом Ек имеет тенденцию к возрастанию, постепенно приближаясь к Ео. Возбудимость при этом растет, так как уменьшается исходная разница между Ео и Ек.
Причиной изменения критического уровня деполяризации под катодом является инактивация натриевой проницаемости, обусловленная длительной деполяризацией мембраны. Вместе с этим значительно повышается проницаемость для К. Все это приводит к тому, что мембрана клетки утрачивает способность отвечать на действие раздражающих стимулов. Такие же изменения мембраны лежат и в основе рассмотренного уже явления аккомодации. Под анодом же при действии тока явления инактивации снижаются .
Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для их изучения нерв или мышцу подвергают действию двух коротких электрических стимулов, следующих друг за другом с определенным интервалом. Первый называется раздражающим, второй — тестирующим. Регистрация возникающих в ответ на эти раздражения ПД позволила установить важные факты.
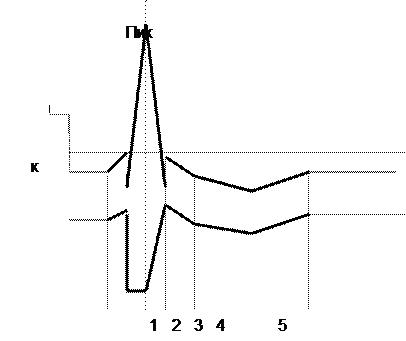 |
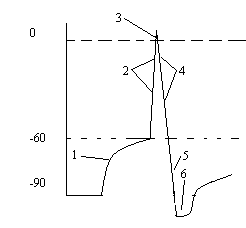
Рисунок 5. Изменения возбудимости при возбуждении.
Обозначения: 1- повышение возбудимости по время локального ответа; 2 – абсолютная рефрактерность; 3- относительная рефрактерность; 4- супернормальная возбудимость во время следовой деполяризации; 5 – субнормальная возбудимость во время следовой гиперполяризации.
Во время локального ответа возбудимость повышена, так как мембрана деполяризована и разность между Е0 и Ек падает. Периоду же возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название абсолютной рефрактерности (невпечатлительности). В это время тестирующий стимул не способен вызвать новый ПД, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности примерно совпадает с длительностью восходящей ветви ПД. В быстро проводящих нервных волокнах она составляет 0,4-0,7 мсек. В волокнах мышцы сердца — 250-300 мсек. Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена.
Согласно ионной теории Ходжкина-Хаксли, абсолютная рефрактерность обусловлена вначале наличием максимальной натриевой проницаемости, когда новый стимул не может что-то изменить или добавить, а затем развитием натриевой инактивации, закрывающей Na-каналы. Вслед за этим происходит снижение натриевой инактивации, в результате чего постепенно восстанавливается способность волокна генерировать ПД. Это — состояние относительной рефрактерности.
Относительная рефрактерная фаза сменяется фазой повышенной (супернормальной) возбудимости, совпадающей по времени с периодом следовой деполяризации. В это время разность между Ео и Ек ниже исходной. В двигательных нервных волокнах теплокровных животных длительность супернормальной фазы составляет 12-30 мсек.
Период повышенной возбудимости сменяется субнормальной фазой , которая совпадает со следовой гиперполяризацией. В это время разница между мембранным потенциалом (Ео) и критическим уровнем деполяризации (Ек) увеличивается. Длительность этой фазы составляет несколько десятков или сотен мсек.
Лабильность. Мы рассмотрели основные механизмы возникновения и распространения в нервных и мышечных волокнах одиночной волны возбуждения. Однако в естественных условиях существования организма по нервным волокнам проходят не одиночные, а ритмические залпы потенциалов действия. В чувствительных нервных окончаниях, расположенных в любых тканях, возникают и распространяются по отходящим от них афферентным нервным волокнам ритмические разряды импульсов даже при очень кратковременном раздражении. Равным образом из ЦНС по эфферентным нервам идет поток импульсов на периферию к исполнительным органам. Если исполнительным органом являются скелетные мышцы, то в них возникают вспышки возбуждений в ритме поступающих по нерву импульсов.
Частота разрядов импульсов в возбудимых тканях может варьировать в широких пределах в зависимости от силы приложенного раздражения, свойств и состояния ткани и от скорости протекания отдельных актов возбуждения в ритмическом ряду. Для характеристики этой скорости Н.Е. Введенским и было сформулировано понятие лабильность. Под лабильностью, или функциональной подвижностью он понимал большую или меньшую скорость протекания тех элементарных реакций, которыми сопровождается возбуждение. Мерой лабильности является наибольшее число потенциалов действия, которое возбудимый субстрат способен воспроизвести в единицу времени в соответствии с частотой подаваемого раздражения.
Первоначально предполагали, что минимальный интервал между импульсами в ритмическом ряду должен соответствовать длительности абсолютного рефрактерного периода. Точные исследования, однако, показали, что при частоте следования стимулов с таким интервалом возникают только два импульса, а третий выпадает вследствие развивающейся депрессии. Поэтому интервал между импульсами должен быть несколько больше абсолютного рефрактерного периода. В двигательных нервных клетках теплокровных животных рефрактерный период составляет около 0,4 мсек, и потенциальный максимальный ритм должен бы быть равным 2500/сек, но на самом деле он равен около 1000/сек. Следует подчеркнуть, что эта частота значительно превышает частоту импульсов, проходящих по этим волокнам в физиологических условиях. Последняя составляет около 100/сек.
Дело в том, что обычно в естественных условиях ткань работает с так называемым оптимальным ритмом. Для пропускания импульсов с таким ритмом не требуется большой силы раздражения. Исследования показали, что частота раздражения и реобаза тока, способного вызывать нервные импульсы с такой частотой, находятся в своеобразной зависимости: реобаза вначале падает по мере роста частоты импульсов, затем снова нарастает. Оптимум находится у нервов в пределах от 75 до 150 имп/сек, для мышц — 20-50 имп/сек. Такой ритм, в отличие от других, очень стойко и длительно может воспроизводиться возбудимыми образованиями.
Таким образом, мы можем теперь назвать все основные параметры возбудимости тканей, характеризующие ее свойства: реобаза, полезное время (хронаксия), критический наклон, лабильность. Все они, кроме последнего, находятся с возбудимостью в обратно пропорциональных отношениях.
Понятие о «парабиозе». Лабильность — величина непостоянная. Она может меняться в зависимости от состояния нерва или мышцы, в зависимости от силы и длительности падающих на них раздражений, от степени утомления и т.п. Впервые изменение лабильности нерва при действии на него сначала химическими, а затем и электрическими раздражителями, изучил Н.Е. Введенский. Он обнаружил закономерное снижение лабильности альтерированного химическим агентом (аммиаком) участка нерва, назвал это явление «парабиозом» и изучил его закономерности. Парабиоз — это обратимое состояние, которое, однако, при углублении действия вызывающего его агента может перейти в необратимое.
Введенский рассматривал парабиоз как особое состояние стойкого не колеблющегося возбуждения, как бы застывшего в одном участке нервного волокна. Действительно, парабиотический участок заряжен отрицательно. Такое явление Введенский считал прообразом перехода возбуждения в торможение в нервных центрах. По его мнению, парабиоз является результатом перевозбуждения нервной клетки слишком сильным или слишком частым раздражением.
Развитие парабиоза протекает в три стадии: уравнительную, парадоксальную и тормозную. Вначале за счет снижения аккомодации отдельные импульсы тока малой частоту при условии их достаточной силы дают уже не 1 импульс, а 2,3 или даже 4. Одновременно порог возбудимости растет, а максимальный ритм возбуждения прогрессивно снижается. В результате на импульсы как малой, так и большой частоты нерв начинает отвечать одной и той же частотой разрядов, наиболее близкой к оптимальному для этого нерва ритму. Это и есть уравнительная фаза парабиоза. На следующем этапе развития процесса в области пороговых интенсивностей раздражения еще сохраняется воспроизведение ритма, близкого к оптимальному, а на частые импульсы ткань или вообще не отвечает, или отвечает очень редкими волнами возбуждения. Это — парадоксальная фаза.
Затем способность волокна к ритмической волновой деятельности падает, падает и амплитуда ПД, увеличивается его длительность., Любое внешнее воздействие подкрепляет состояние торможения нервного волокна и одновременно затормаживается само. Это — последняя, тормозная фаза парабиоза.
В настоящее время описанный феномен объясняется с позиций мембранной теории нарушением механизма повышения натриевой проницаемости и появлением затяжной натриевой инактивации. В результате этого Na-каналы остаются закрытыми, он накапливается в клетке и наружная поверхность мембраны длительное время сохраняет отрицательный заряд. Это препятствует новому раздражению за счет удлинения рефрактерного периода. При набегании на участок парабиоза часто следующих друг за другом ПД, инактивация натриевой проницаемости, вызванная альтерирующим агентом, суммируется с инактивацией, сопровождающей нервный импульс. В результате возбудимость снижается настолько, что проведение следующего импульса полностью блокируется.
Обмен веществ и энергии при возбуждении. При возникновении и проведении возбуждения в нервных клетках и мышечных волокнах происходит усиление обмена веществ. Это проявляется как в ряде биохимических изменений, происходящих в мембране и протоплазме клеток, так и в усилении их теплопродукции. Установлено, что при возбуждении происходят: усиление распада в клетках богатых энергией соединений — АТФ и креатинфосфата (КФ), усиление процессов распада и синтеза углеводов, белков и липидов, усиление окислительных процессов, приводящих в сочетании с гликолизом к резинтезу АТФ и КФ, синтез и разрушение ацетилхолина и норадреналина, других медиаторов, усиление синтеза РНК и белков. Все эти процессы более всего выражены в период восстановления состояния мембраны после ПД.
В нервах и мышцах каждая волна возбуждения сопровождается выделением двух порций тепла, из которых первая называется начальным, а вторая — запаздывающим теплом. Начальное теплообразование происходит в момент возбуждения и составляет незначительную часть всей теплопродукции (2-10%) при возбуждении. Предполагают, что это тепло связано с теми физико-химическими процессами, которые развиваются в момент генерации ПД. Запаздывающее теплообразование происходит в течение более продолжительного времени, и длится многие минуты. Оно связано с теми химическими процессами, которые происходят в ткани вслед за волной возбуждения, и , по образному выражению Ухтомского, составляют «метаболический хвост кометы возбуждения».
Изменения возбудимости при возбуждении. Нормальная физиология
Изменения возбудимости при возбуждении
При развитии потенциала действия происходят фазные изменения возбудимости ткани (рис. 2). Состоянию исходной поляризации мембраны (мембранный потенциал покоя) соответствует нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена. Эта фаза возбудимости получила название повышенной возбудимости (первичной экзальтации). В это время мембранный потенциал приближается к критическому уровню деполяризации, поэтому дополнительный стимул, даже если он меньше порогового, может довести мембрану до критического уровня деполяризации. В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны и возникает в связи с тем, что натриевые каналы инактивируются.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня – фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны, достигая величины критического уровня деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя.
Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной экзальтации). Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации по сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен и новое возбуждение может возникнуть при действии раздражителей подпороговой силы.
В период развития положительного следового потенциала возбудимость ткани понижена – фаза субнормальной воз
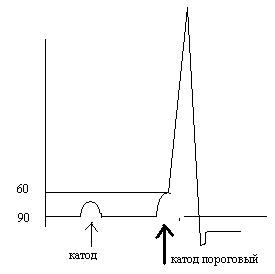
Поделитесь на страничке
Следующая глава >
3.7.Изменения возбудимости клетки во время ее возбуждения. Лабильность
А. Возбудимость
клетки во время ее возбуждения быстро
и сильно изменяется.
Различают несколько фаз изменения
возбудимости, каждая из которых строго
соответствует определенной фазе ПД и,
так же как и фазы ПД, определяется
состоянием проницаемости клеточной
мембраны для ионов. Схематично эти
изменения представлены на рис.
3.6.б.
1.
Кратковременное
повышение возбудимости
в начале развития ПД, когда уже возникла
частичная деполяризация клеточной
мембраны. Если деполяризация не достигает
критической величины, то регистрируется
локальный потенциал. В случае, если
деполяризация достигает Екр, то
развивается ПД. При замедленном развитии
начальной деполяризации она оценивается
как препотенциал. Возбудимость повышена
потому, что клетка частично деполяризована,
мембранный потенциал приближается к
критическому уровню, поскольку
открывается часть потенциалчувствительных
быстрых
Na-каналов.
При этом достаточно небольшого увеличения
силы раздражителя, чтобы деполяризация
достигла Екр,
при которой возникает ПД.
2.
Абсолютная
рефракторная фаза
— это
полная невозбудимость клетки (возбудимость
равна нулю), она соответствует пику ПД
и продолжается
1-2 мс; если
ПД более продолжителен, то более
продолжительна и абсолютная рефракторная
фаза. Клетка в этот период при любой
силе раздражения не отвечает.
Невозбудимость клетки в фазу деполяризации
и инверсии (в первую ее половину
— восходящая
часть пика ПД) объясняется тем, что
потенциалзависимые т-ворота
Na-каналов
уже открыты и ионы
Na+ быстро
поступают в клетку по всем каналам. Те
ворота
Na-каналов,
которые еще не успели открыться,
открываются под влиянием деполяризации
— уменьшения
мембранного потенциала. Поэтому
дополнительное раздражение клетки
относительно движения ионов
Na+
в клетку ничего изменить не может.


Рис.
3.6.
Фазовые изменения возбудимости клетки
(б)
во время ПД (а). 1,4
—
возбудимость повышена;
2 —
абсолютная рефрактерная фаза;
3 —
относительная рефрактерная фаза
Именно поэтому
ПД либо совсем не возникает на раздражение,
если оно мало, либо возникает максимальным,
если оно достаточной силы (пороговой
или сверхпороговой). В период нисходящей
части фазы инверсии и реполяризации
клетка невозбудима потому, что закрываются
инактивационные
h-ворота Na-каналов,
в результате чего клеточная мембрана
непроницаема для иона
Na+
даже при сильном раздражении. Кроме
того, в этот период открываются уже в
большом количестве К-каналы,
К+
быстро выходит из клетки, обеспечивая
нисходящую часть фазы инверсии и
реполяризацию. Абсолютная рефрактерная
фаза в процессе реполяризации продолжается
до момента, когда мембранный потенциал
будет примерно на уровне Екр.
В это время около половины Nа+
—каналов
возвращается в исходное состояние,
поэтому возможна их новая активация.
Абсолютный рефракторный период
ограничивает максимальную частоту
генерации ПД. Если абсолютный рефракторный
период завершается через
2 мс после
начала ПД, то клетка может возбуждаться
с частотой максимум
500 имп/с.
Существуют клетки с еще более коротким
рефракторным периодом, в которых
возбуждение может в крайних случаях
повторяться с частотой
1000 имп/с.
Такие клетки встречаются в ретикулярной
формации ЦНС.
3.
Относительная
рефракторная фаза
— это
период восстановления возбудимости,
когда сильное раздражение может вызвать
новое возбуждение (см. рис.
3.6,6,
кривая
3).
Относительная рефрактерная фаза
соответствует конечной части фазы
реполяризации от уровня Екр ±
10 мВ и
следовой, гиперполяризации клеточной
мембраны, что является следствием все
еще повышенной проницаемости для ионов
К+
и избыточного выхода ионов К+-каналов
из клетки. Поэтому, чтобы вызвать
возбуждение в этот период, необходимо
приложить более сильное раздражение,
так как часть Nа+
— каналов
в конце реполяризации находится еще в
состоянии инактивации, а выход ионов
К+
из клетки препятствует ее деполяризации.
Кроме того, в период следовой
гиперполяризации мембранный потенциал
больше и, естественно, дальше отстоит
от критического уровня деполяризации.
Если реполяризация в конце пика ПД
замедляется (см. рис. 3.6,а), то относительная
рефрактерная фаза включает и период
замедления реполяризации, и период
гиперполяризации.
4.
Фаза
экзальтации
— это
период повышенной возбудимости. Он
соответствует следовой деполяризации.
В нейронах ЦНС вслед за гиперполяризацией
возможна частичная деполяризация
клеточной мембраны. В эту фазу очередной
ПД можно вызвать более слабым раздражением,
поскольку мембранный потенциал несколько
ниже обычного и оказывается ближе к
критическому уровню деполяризации,
что объясняют повышенной проницаемостью
клеточной мембраны для ионов
Na+.
Скорость протекания фазовых изменений
возбудимости клетки определяет ее
лабильность.
Б. Лабильность,
или функциональная подвижность
(Н.Е.Введенский)
— это
скорость протекания одного цикла
возбуждения, т.е. ПД. Как видно из
определения, лабильность ткани зависит
от длительности ПД. Это означает, что
лабильность, как и ПД, определяется
скоростью перемещения ионов в клетку
и из клетки, которая, в свою очередь,
зависит от скорости изменения
проницаемости клеточной мембраны.
Особое значение при этом имеет
длительность рефракторной фазы: чем
больше рефракторная фаза, тем ниже
лабильность ткани.
Мерой лабильности
является максимальное число ПД, которое
ткань может воспроизвести в
1 с. В
эксперименте лабильность исследуют в
процессе регистрации максимального
числа ПД, которое может воспроизвести
клетка при увеличении частоты ритмического
раздражения.
Лабильность
различных клеток существенно различается.
Так, лабильность нерва равна
500-1000,
нейронов
— 20-200,
синапса
— порядка
100 импульсов
в секунду. Лабильность клеток понижается
при длительном бездействии и при
утомлении.
Следует отметить,
что при постепенном увеличении частоты
ритмического раздражения лабильность
ткани повышается, т.е. ткань отвечает
более высокой частотой возбуждения по
сравнению с исходной частотой. Это
явление открыто А.А.Ухтомским и называется
усвоением ритма раздражения.
Фазы возбудимости при возбуждении.
При развитии потенциала действия
происходит изменение возбудимости
ткани, причем, это изменение протекает
по фазам (рис.2).
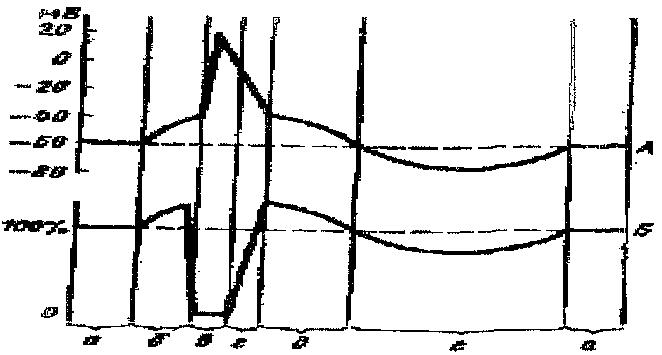
Рис. 2. Соотношение
одиночного цикла возбуждения (А) и фаз
возбудимости
(Б).
А: а —
мембранный потенциал покоя; б — предспайк,
локальный ответ, ВПСП; в —
спайк,
потенциал действия, деполяризация и
инверсия; г — потенциал действия,
реполяризации; д
—
отрицательный следовой потенциал,
следовая деполяризация; е – положительный
следовой
потенциал, следовая гиперполяризация.
Б: а — исходный уровень возбудимости;
б — фаза первичной экзальтации, повышенная
возбудимость; в — фаза абсолютной
рефрактерности;
г — фаза относительной рефрактерности;
д — фаза вторичной . ; е
— фаза вторичной рефрактерности.
Состоянию исходной поляризации
мембраны, которую отражает мембранный
потенциал покоя, соответствует исходное
состояние ее возбудимости и,
следовательно, клетки — это нормальный
уровень возбудимости. В период локального
ответа возбудимость ткани повышена,
эта фаза возбудимости получила
название первичной
экзальтации. Во
время развития локального ответа
мембранный потенциал покоя приближается
к критическому уровню деполяризации
и для достижения последнего достаточна
сила раздражителя меньшая, чем
пороговая (подпороговая).
В период развития пикового
потенциала идет лавинообразное
поступление ионов натрия внутрь
клетки, в результате чего происходит
перезарядка мембраны и она утрачивает
способность отвечать возбуждением на
раздражители даже сверхпороговой силы.
Эта фаза возбудимости получила
название абсолютной
рефрактерности
(абсолютной
невозбудимости). Ома длится до конца
перезарядки мембраны. Абсолютная
рефрактерность, т. е. полная не возбудимость
мембраны возникает в связи с тем, что
натриевые каналы в начале полностью
открываются, а затем инактивируются.
После окончания фазы
перезарядки мембраны возбудимость ее
постепенно восстанавливается до
исходного уровня — фаза
относительной рефрактерности.
Она продолжается до
восстановления заряда мембраны до
величины, соответствующей критическому
уровню деполяризации. Так как и этот
период мембранный потенциал покоя еще
не восстановлен, то возбудимость ткани
понижена, и новое возбуждение может
возникнуть только при действии
сверхпорогового раздражителя. Снижение
возбудимости в фазу относительной
рефрактерности связано с частичной
инактивацией натриевых каналов и
активацией калиевых.
Периоду отрицательного
следового потенциала соответствует
повышенный уровень возбудимости — фаза
вторичной экзальтации.
Так как мембранный
потенциал в эту фазу ближе к критическому
уровню деполяризации, но сравнению
с состоянием покоя (исходной поляризацией),
то порог раздражения снижен, т. е.
возбудимость повышена. В эту фазу новое
возбуждение может возникнуть при
действии раздражителей подпороговой
силы. Натриевые каналы в эту фазу
инактивированы неполностью. В Период
развития положительного следового
потенциала возбудимость ткачи
понижена — фаза вторичной
рефрактерности.
В эту фазу мембранный
Потенциал увеличивается (состояние
гиперполяризации мембраны), удаляясь
от критического уровня деполяризации,
порог раздражения повышается и новое
возбуждение может возникнуть только
при действии раздражителей
сверхпороговой величины. Гиперполяризация
мембраны развивается вследствие
трех причин: во-первых, продолжающимся
выходом ионов калия; во-вторых, открытием,
возможно, каналов для хлора и поступление
этиx
ионов в цитоплазму клетки; в-третьих,
усиленной работой натрий-калиевого
насоса.
Закон силы-длительности:
раздражающее действие постоянного тока
зависит не только от его величины, но и
от времени, в течение которого он
Действует. Чем больше ток, тем меньше
времени он должен действовать для
возникновения возбуждения.
Исследования зависимости силы-длительности
показали, что последняя имеет
гиперболический характер (рис. 3).
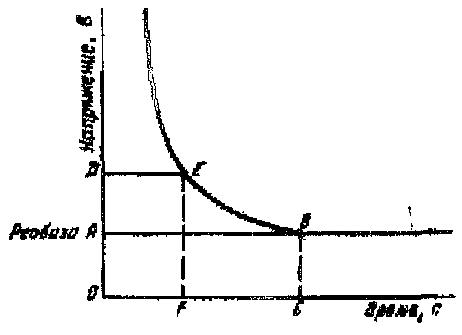
Рис3.
Графическое выражение законы
силы-длительности.
Из этого следует, что сок
ниже некоторой минимальной величины
не вызывает возбуждение, как бы длительно
он не действовал, и чем короче импульсы
тока, тем меньшую раздражающую
способность они имеют. Причиной такой’
зависимости является мембранная
емкость. Очень «короткие» токи
просто не успевают разрядить эту
емкость до критического уровня
деполяризации. Минимальная величина
тока, способная вызвать возбуждение
при неограниченно длительном его
действии, называется реобазой.
Время, в течение
которого действует ток, равный
реобазе, и вызывает возбуждение,
называется полезным
временем.
В связи с тем, что определение
этого времени затруднено, было введено
понятие хронаксия —
минимальное время, в
течение которого ток, равный двум
реобазам, должен действовать на ткань,
чтобы вызвать ответную реакцию.
Определение хронаксии — хронаксиметрия
— находит применение
в клинике. Электрический ток, приложенный
к мышце, проходит через как мышечные,
так и нервные волокна, и их окончания,
находящиеся в этой мышце. Так как
хронаксия нервных волокон значительно
меньше хронаксии мышечных волокон,
то при исследовании хронаксии мышцы
практически получают хронаксию
нервных волокон. Если нерв поврежден
или произошла гибель соответствующих
мотонейронов спинного мозга (это имеет
место при полимиелите и некоторых других
заболеваниях), то происходит
перерождение нервных волокон и тогда
определяется хронаксия уже мышечных
волокон, которая имеет большую величину,
чем нервных волокон.
Изменения возбудимости скелетной мышцы во время одиночного цикла возбуждения.
Изменение возбудимости протекает по фазам, которые зависят от фаз потенциала действия
фазы возбудимости:
1. Фаза первичной экзальтации:Возникает в начале возбуждения, когда мембранный потенциал изменяется до критического уровня.Соответствует латентному периоду потенциала действия (периоду медленной деполяризации). Характеризуется незначительным повышением возбудимости
2. Фаза абсолютной рефрактерности: Совпадает с восходящей частью пикового потенциала, когда мембранный потенциал изменяется от критического уровня до «спайка». Соответствует периоду быстрой деполяризации. Характеризуется полной невозбудимостью мембраны (даже самый большой по силе раздражитель не вызывает возбуждение)
3. Фаза относительной рефрактерности:Совпадает с нисходящей частью пикового потенциала, когда мембранный потенциал изменяется от «спайка» к критическому уровню, оставаясь выше него. Соответствует периоду быстрой реполяризации. характеризуется пониженной возбудимостью (возбудимость постепенно увеличивается, но остается ниже, чем в состоянии покоя).В этот период может возникнуть новое возбуждение, но сила раздражителя должна превыышать пороговую величину
4. Фаза вторичной экзальтации (супернормальной возбудимости):Возникает в конце возбуждения, когда мембранный потенциал, минуя критический уровень, изменяется до величины потенциала покоя. Соответствует периоду следовой деполяризации. Характеризуется повышенной возбудимостью (мембрана может ответить новым возбуждением даже на действие подпорогового раздражителя)
5. Фаза субнормальной возбудимости:Возникает в конце возбуждения, когда изменение мембранного потенциала происходит ниже уровня потенциала покоя. Соответствует периоду следовой гиперполяризации. характеризуется пониженной возбудимостью

Физиология возбудимых тканей — Знаешь как
Содержание статьи
 Раздражимость
Раздражимость
Раздражимостью называют способность живых систем под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности. Формы проявления раздражимости весьма разнообразны. У одних клеток при этом изменяются форма, структура, их рост и процесс деления; в других образуются различные химические соединения, совершается та или иная работа, движение.
Раздражителем живой клетки или организма как целого может оказаться любое изменение внешней среды или внутреннего состояния организма, если оно достигает определенной величины.
Среди раздражителей различают физические (температура, укол, давление, свет, звук, электрические раздражители), физико-химические (изменения осмотического давления, активной реакции среды, электролитного состава, коллоидного состояния) и химические (химические вещества пищи, химические соединения, образующиеся в организме,— гормоны, продукты обмена веществ и т. п.).
Рис. 17. Первый (I) и второй (II) опыты Гальвани
Естественными раздражителями клеток, вызывающими их деятельность, являются нервные импульсы.
Возбудимость
Клетки нервной и мышечной тканей приспособлены к осуществлению быстрых реакций на раздражение. Клетки этих тканей называют возбудимыми, а их способность отвечать на раздражение возбуждением называют возбудимостью.
Возбуждение способно перемещаться из одного места клетки в другое, из одной клетки в другую.
 Рис. 18. Последовательные этапы приготовления нервно-мышечного препарата.
Рис. 18. Последовательные этапы приготовления нервно-мышечного препарата.
Возбуждение характеризуется комплексом химических, функциональных, физико-химических, электрических явлений. Обязательным признаком возбуждения является изменение электрического состояния поверхностной клеточной мембраны. Именно электрические явления обеспечивают проведение возбуждения в возбудимых тканях.
Биоэлектрические явления
Возникновение и распространение возбуждения связано с изменением электрического заряда живой ткани, с так называемыми биоэлектрическими явлениями.
Электрические явления у животных известны давно. Еще в 1776 г. они были описаны у электрического ската. Началом же экспериментального изучения электрических явлений в животных тканях следует считать опыты итальянского врача Луиджи Гальвани (1791). В опытах он использовал препараты задних конечностей лягушки, соединенных с позвоночником. Подвешивая эти препараты на медном крючке к железным перилам балкона, он обратил внимание, что, когда конечности лягушки раскачивались ветром, их мышцы сокращались при каждом прикосновении к перилам. На основании этого Гальвани пришел к выводу, что подергивания лапок были вызваны «животным электричеством», зарождающимся в спинном мозге лягушки и передаваемым по металлическим проводникам (крючку и перилам балкона) к мышцам препарата.
 Рис. 19. Схема вторичного сокращения.
Рис. 19. Схема вторичного сокращения.
Против этого положения Гальвани о «животном электричестве» выступил физик Александр Вольта. В 1792 г. Вольта повторил опыты Гальвани и установил, что описанные Гальвани явления нельзя считать «животным электричеством». В опыте Гальвани источником тока служил не спинной мозг лягушки, а цепь, образованная из разнородных металлов — меди и железа.
Вольта был прав. Первый опыт Гальвани не доказывал наличия «животного электричества», но эти исследования привлекли внимание ученых к изучению электрических явлений в живых образованиях.
В ответ на возражение Вольта Гальвани произвел второй опыт, уже без участия металлов. При этом также наблюдалось сокращение лапки лягушки.
Опыт Гальвани. Для этого обездвижьте лягушку и перережьте ее поперек в области верхних грудных позвонков. Захватив остаток позвоночника салфеточкой, снимите с задних конечностей кожу, а затем пинцетом удалите остатки внутренностей. Становятся хорошо видны нервные стволики крестцового сплетения, расположенные с обеих сторон позвоночника пучками. Подведите под оба пучка нервных волокон одну пластинку пинцета Гальвани, а другой пластинкой пинцета прикоснитесь к нервам сверху. Мышцы лапок при этом сокращаются (рис. 17, I). Пинцет Гальвани состоит из цинковой и медной пластинок. Объясните, почему сокращаются мышцы лапок в опыте Гальвани.
 Рис. 20. Стеклянные микроэлектроды:
Рис. 20. Стеклянные микроэлектроды:
А — одноканальный; Б — двухканальный; справа — кончик электрода около тела нейрона
А теперь приготовьте нервно-мышечный препарат лягушки.
Основные этапы приготовления нервно-мышечного препарата приведены на рисунке 18.
Лягушку обездвижьте. Возьмите ее левой рукой за бедра (в этом положении хорошо выделяется позвоночник) и перережьте позвоночник на 1—1,5 см выше места отхождения тазовых костей (рис. 18, 1). Свисающую переднюю часть туловища и внутренности удалите. Остаток позвоночника крепко держите пинцетом или левой рукой. Другим пинцетом захватите кожу около позвоночника и тяните ее вниз, чтобы, выворачивая, снять с конечностей (рис. 18, 2). Конечности положите на чистую тарелку и залейте раствором Рингера. Руки вымойте или тщательно вытрите от слизи, покрывающей кожу лягушки. Захватите пинцетом или рукой кусочек позвоночника и подогните его вниз так, чтобы конечности висели под углом к позвоночнику и хорошо выделялась копчиковая кость (рис. 18, 3).
Осторожно вырежьте копчиковую кость. Ножницы при этом держите как можно ближе к кости, чтобы не повредить идущие параллельно с обеих сторон нервы. Вырезав копчик, положите препарат на тарелку и разделите его на две половины. Для этого перережьте вдоль сначала остаток позвоночника, а затем лобковое сочленение (рис. 18, 4).
 Рис. 21. Схема измерения электрической активности мышечного волокна А с помощью микроэлектрода:
Рис. 21. Схема измерения электрической активности мышечного волокна А с помощью микроэлектрода:
М — микроэлектрод; И — индифферентный электрод. Луч на экране осциллографа Б показывает, что до прокола микроэлектродом мембраны разность потенциалов между М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена электроотрицательно по отношению к ее наружной поверхности.
Одну конечность оставьте как запасную, сохраняя ее в растворе Рингера; другую положите на спинную сторону и отделите ножницами подвздошную кость. Захватив пинцетом кусочек позвоночника, отведите в сторону седалищный нерв и удалите подвздошную кость. При помощи двух пинцетов раздвиньте мышцу на спинной поверхности бедра по средней линии (рис. 18, 5). Осторожно не касаясь ножницами и пинцетом нерва, отделите его от окружающих тканей, вдоль бедра до колен. (Лучше это делать стеклянным крючком.) Нерв отведите в сторону и освободите бедренную кость от мышц (рис. 18, б). На голени отделите от кости икроножную мышцу, подрезав ахиллово сухожилие, и привяжите к нему нитку. Голень и стопу отрежьте ниже колена (рис. 18, 7). Препарат положите в стакан с раствором Рингера.
Проделайте второй опыт Гальвани (сокращение без металла). Для этого нервно-мышечный препарат положите на дощечку. Отрежьте кусочек мышцы и стеклянным крючком быстро набросьте нерв препарата на пораненный участок мышцы так, чтобы он одновременно коснулся поврежденной и неповрежденной поверхности мышцы (рис. 17, II). Мышца при этом сокращается. Объясните, почему это происходит.
Гальвани все же оказался прав в своем утверждении о существовании «животного электричества», что позже было подтверждено исследованиями других ученых.
В этом отношении интересны опыты Маттеучю, получившие название вторичного сокращения.
На мышцу одного нервно-мышечного препарата набросьте нерв другого нервно-мышечного препарата (рис. 19) и раздражайте электрическим током нерв первого препарата. Вы наблюдаете сокращение мышцы и второго препарата. Это объясняется тем, что при возбуждении в мышце первого препарата возникают токи действия, которые вызывают возбуждение второго нервно-мышечного препарата.
В дальнейшем в изучение биоэлектрических явлений очень важный вклад внесли русские ученые, среди них И. М. Сеченов, обнаруживший с помощью гальванометра электрические явления в головном мозге, Н. Е. Введенский, А. Ф. Самойлов и др.
В настоящее время имеются весьма совершенные, высокочувствительные приборы (электронно-лучевые трубки с электронными усилителями), позволяющие регистрировать электрические явления в тканях и органах.
Потенциал покоя и потенциал действия мышц
Между наружной поверхностью клетки и ее цитоплазмой в состоянии покоя существует разность потенциалов около 60—90 мВ, причем поверхность клетки заряжена электроположительно по отношению к цитоплазме. Эту разность потенциалов называют потенциалом покоя или мембранным потенциалом. Точное измерение потенциала покоя возможно только с помощью микроэлектродов, предназначенных для внутриклеточного отведения токов, очень мощных усилителей и чувствительных регистрирующих приборов — осциллографов.
 Рис. 22. Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Рис. 22. Потенциал действия аксона кальмара, отводимый с помощью внутриклеточного электрода.
Микроэлектрод (рис. 20) представляет собой тонкий стеклянный капилляр, кончик которого имеет диаметр около микрона. Этот капилляр заполняют солевым раствором, погружают в него металлический электрод и соединяют с усилителем и осциллографом (рис. 21). Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа отклоняется вниз из своего исходного положения и устанавливается на новом уровне. Это свидетельствует о наличии разности потенциалов между наружной и внутренней поверхностью клеточной мембраны.
Наиболее полно происхождение потенциала покоя объясняет так называемая мембранно-ионная теория. Согласно этой теории все клетки покрыты мембраной, имеющей неодинаковую проницаемость для различных ионов. В связи с этим внутри клетки в цитоплазме в 30—50 раз больше ионов калия, в 8—10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем на поверхности клетки. В состоянии покоя клеточная мембрана более проницаема для ионов калия, чем для ионов натрия.
Диффузия положительно заряженных ионов калия из цитоплазмы на поверхность клетки придает наружной поверхности мембраны положительный заряд.
Таким образом, поверхность клетки в покое несет на себе положительный заряд, тогда как внутренняя сторона мембраны оказывается заряженной отрицательно за счет ионов хлора, аминокислот и других крупных органических анионов, которые через мембрану практически не проникают.
Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя, то в этом участке возникает возбуждение, проявляющееся в быстром колебании мембранного потенциала и называемое потенциалом действия.
Потенциал действия можно зарегистрировать с помощью электродов, приложенных к внешней поверхности волокна (внеклеточное отведение), либо микроэлектрода, введенного в цитоплазму (внутриклеточное отведение) (рис. 22).
При введении микроэлектрода в нервное волокно сначала регистрируется потенциал величиной около 60 мВ. Это мембранный потенциал.
Потенциал действия представляет собой резкий скачок мембранного потенциала. При этом происходит не только изменение отрицательного потенциала покоя от величины — 60 мВ до нуля, но и переход за нулевую линию на несколько десятков милливольт, так что амплитуда потенциала действия превышает потенциал покоя и составляет приблизительно 90 мВ.
В потенциале действия различают его пик (спайк, по терминологии английских авторов) и следовые потенциалы. Во время пика мембранный потенциал меняет свой знак.
Причина возникновения потенциала действия — изменение ионной проницаемости мембраны. При раздражении проницаемость клеточной мембраны для ионов натрия повышается. Натрий стремится внутрь клетки, так как, во-первых, он заряжен положительно и его влекут внутрь электростатические силы, вовторых, концентрация его внутри клетки невелика. В покое клеточная мембрана была малопроницаемой для этого иона. Раздражение изменило проницаемость мембраны, и поток положительно заряженных ионов натрия из внешней среды клетки в цитоплазму значительно превысил поток ионов калия из клетки наружу. В результате произошло извращение потенциала мембраны (фаза деполяризации). Внутренняя поверхность мембраны стала заряженной положительно, а наружная вследствие потери положительно заряженных ионов натрия — отрицательно. В этот момент и регистрируется пик потенциала действия.
Повышение проницаемости мембраны для ионов натрия продолжается очень короткое время. Вслед за этим в клетке возникают восстановительные процессы, приводящие к тому, что проницаемость мембраны для ионов натрия вновь понижается, а для ионов калия возрастает. Поскольку ионы калия также заряжены положительно, то, выходя из клетки, они восстанавливают исходные отношения снаружи и внутри клетки (фаза реполяризации).
Накопления натрия внутри клетки при многократном возбуждении ее не происходит, потому что натрий эвакуируется из нее постоянно за счет действия специального биохимического механизма, называемого «натриевым насосом». Есть данные и об активном транспорте калия с помощью «натрий-калиевого насоса».
Таким образом, согласно мембранной теории в происхождении биоэлектрических явлений решающее значение имеет избирательная проницаемость клеточной мембраны, обусловливающая разный ионный состав на поверхности и внутри клетки, а следовательно, и разный заряд этих поверхностей. Следует заметить, что многие положения мембранной теории все еще дискуссионны и нуждаются в углубленной разработке.
Итак, биоэлектрические свойства клеток, обусловленные неравномерным распределением минеральных ионов, играют ведущую роль в процессах клеточного возбуждения. Возникновение потенциала действия является наиболее характерным показателем возбуждения в клетках, тканях и органах. Поэтому электрофизиологические исследования широко применяются в экспериментальных лабораториях и в клиниках.
Значение регистрации биоэлектрических явлений. Регистрация электрических потенциалов в живом организме дает возможность судить об изменениях многих физиологических функций. Так как электрическая активность многих органов человека в норме имеет типичные и постоянные характеристики, то методы электрофизиологии широко используются для диагностики болезней в практической медицине. Большие успехи достигнуты в тонком распознавании болезней сердца, нервной системы, мышц. Электрофизиологические методы сыграли важную роль в решении многих проблем космической физиологии. С помощью методов телеметрии оказалось возможным передавать информацию о состоянии сердечной мышцы, деятельности мозга, скелетной мускулатуры и других органов в условиях невесомости, перегрузок.
Изучение электрической активности центральной нервной системы легло в основу конструирования сложных счетно-решающих электронных машин.
Под электроэнцефалографией понимают запись биоэлектрических явлений, протекающих в головном мозге, преимущественно в коре больших полушарий головного мозга.
Для отведения биотоков от различных структур головного мозга используют различные конструкции электродов. В эксперименте на животных электроды можно ввести через кости черепа прямо в нужный участок головного мозга. Такие «вживленные» электроды долго удерживаются в мозгу специальным креплением и позволяют изучать электрическую активность определенных участков головного мозга при различных состояниях животного.
 Рис. 23. Шлемы для крепления электродов.
Рис. 23. Шлемы для крепления электродов.
В настоящее время возможна длительная регистрация электрических явлений даже в отдельных клетках мозга с помощью микроэлектродов.
При записи биотоков мозга человека — электроэнцефалограммы — пользуются обычно серебряными электродами, имеющими вид пластинки размером с двухкопеечную монету. Электроды на голове испытуемого человека укрепляются с помощью шлемов-сеток. Шлемы изготовляют из эластичных резиновых тяжей, натяжение которых регулируют. Шлемы, плотно прилегая к голове испытуемого, надежно удерживают электроды (рис. 23).
Запись биотоков мозга производится на приборах — электроэнцефалографах, имеющих разную конструкцию и включающих несколько усилителей биотоков, осциллографы и сложный пульт управления ими.
В настоящее время выпускают приборы, позволяющие регистрировать одновременно электрическую активность от 2 до 32 точек мозга и более.
Для точного анализа электрических явлений, возникающих в головном мозге и других органах, используют электронно-вычислительные машины.
Электрическая активность мозга человека носит ритмический характер. Электроды, расположенные на поверхности головы, дают возможность зарегистрировать биотоки сразу от многих клеток мозга, лежащих под ними. Поэтому общий характер энцефалограммы оказывается очень сложным. Вместе с тем удалось установить, что наиболее выраженных и часто встречающихся ритмов колебания электрической активности немного. Названия этих ритмов условно обозначены греческими буквами α, β, δ, ϑ (рис. 25, табл. 4).
Таблица 4
Ритмы электроэцефалограммы
| Название ритма | Частота колебаний в 1 с |
| Дельта-ритм Тета-ритм Альфа-ритм Бета-ритм | 0,5— 3 4— 7 8—13 14—35 |
 Рис. 25. Основные ритмы коры мозга человека:
Рис. 25. Основные ритмы коры мозга человека:
1 —альфа-ритм- 2 — бета-ритм; 3 — дельта-ритм; 4-тета-ритм; 5-веретена; вверх у — отметка времени
При ограниченном поступлении центростремительных импульсов в исследуемый участок мозга обычно наблюдаются медленные волны с большим размахом колебаний. Если в кору поступают многочисленные импульсы, то клетки в этом участке могут находиться в разных стадиях возбуждения и общая электрическая активность над этим участком характеризуется частыми колебаниями с небольшой амплитудой типа бета-ритма. На рисунке 26 приведена электроэнцефалограмма с затылочной области коры (там располагается зрительная зона коры больших полушарий). На ней отчетливо виден переход альфа-ритма в бета-ритм и обратно при открывании и закрывании глаз, т. е. при увеличении и снижении потока центростремительных импульсов к зрительной зоне коры.
Современная техника позволяет регистрировать энцефалограмму у человека на расстоянии и даже в условиях космического полета.
Получил достаточно широкое распространение и метод исследования электрической активности мышц — электромиография.
 Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальная паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Для отведения биопотенциалов мышц человека используют накожные металлические электроды диаметром 10 мм. Электроды укрепляют на исследуемой мышце эластической манжетой; между ними и кожей находится обычно специальная паста, улучшающая контакт с телом и электропроводность. Колебания биопотенциалов мышц имеют также ритмический характер, только частота их и амплитуда значительно больше, чем при записи электроэнцефалограммы. Усиление мышечной активности сопровождается увеличением амплитуды и колебаний электромиограммы. При утомлении мышц частота колебаний, как правило, падает.
Рис. 26. Изменения электроэнцефалограммы затылочной области коры, показывающие переход от альфа-ритма к бета-ритму при открывании глаз (стрелка вверх) и восстановление альфа-ритма при закрывании глаз (стрелка вниз).
На рисунке 27 представлены электромиограммы трехглавой и двуглавой мышц плеча при работе напильником. На них отчетливо видно усиление колебаний при возбуждении мышц. Электромиография позволяет установить степень участия тех или иных мышц в выполняемом движении. Особенно важен этот метод для установления наличия паралича мышц при некоторых заболеваниях. Так, с помощью электромиографии можно обнаружить паралич дыхательных мышц при полиомиелите до того, как остановится дыхание. А это очень важно для принятия необходимых мер (перевод на управляемое дыхание с помощью специальной аппаратуры) с целью спасения жизни человеку.
 и двуглавой (2) мышц плеча") Рис. 27. Электромиограммы трехглавой (1) и двуглавой (2) мышц плеча у рабочего при опиловке.
Рис. 27. Электромиограммы трехглавой (1) и двуглавой (2) мышц плеча у рабочего при опиловке.
Данные электромиографии используются при создании активных протезов и управляемых манипуляторов.
«Искусственная рука» — одно из первых устройств, управляемых с помощью биотоков мышц. Это активный протез, воспроизводящий нормальные движения кисти. Такой активный протез имеет браслет для отведения биотоков мышц сохранившейся культи. На браслете укреплены металлические чашечки, заполненные токопроводящей пастой. Отводимые с помощью браслета мышечные токи усиливаются и поступают в специальный блок на протезе, где формируется управляющий сигнал. Сигнал приводит в действие миниатюрный двигатель, который обеспечивает движение протеза. Сам протез выполняется из пластических материалов, сходных по форме, размерам и цвету с нормальной рукой.
Передача возбуждения в синапсах
Возбуждение от одной нервной клетки к другой или от нервного волокна к мышечной или железистой клетке передается с помощью синапсов.
Аксоны большинства нейронов, подходя к другим нервным клеткам, ветвятся и образуют многочисленные окончания на телах этих клеток и их дендринах (рис. 28). Такие места контактов называют синапсами.
 Рис. 28. Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Рис. 28. Расположение синапсов на теле нейрона и на его дендритах (по Хаачу и Барру)
Количество синапсов на теле одного нейрона достигает 100 и больше, а на дендритах одного нейрона — нескольких тысяч. Одно нервное волокно может образовать до 10 000 синапсов на многих нервных клетках.
Синапс имеет сложное строение (рис. 29). Он образован двумя мембранами — пресинаптической и постсинаптической, между которыми есть синоптическая щель. Пресинаптическая мембрана находится на нервном окончании. Нервные окончания в центральной нервной системе имеют вид пуговок, колечек или бляшек. На теле пирамидной клетки насчитывается несколько тысяч нервных окончаний. Каждая синаптическая пуговка покрыта пресинаптической мембраной. Постсинаптическая мембрана находится на теле или дендритах нейрона, к которому передается нервный импульс. В пресинаптической области обычно наблюдаются большие скопления митохондрий.
Возбуждение через синапсы передается химическим путем с помощью особого вещества — посредника, или медиатора, находящегося в синаптической бляшке. В синаптической бляшке медиатор находится в синаптических пузырьках, каждый из которых содержит до 2000 молекул медиатора. В разных синапсах вырабатываются разные медиаторы. Чаще всего в качестве медиатора служит ацетилхолин, адреналин или норадреналин в некоторых синапсах — глютаминовая кислота.
Каков же механизм передачи возбуждения в синапсах? Приход нервного импульса в пресинаптическое окончание сопровождается синхронным выбросом в синаптическую щель медиатора из синаптических пузырьков, расположенных в непосредственной близости от нее. Размеры этой щели очень малы (около 200 Å), и медиатор, быстро достигая постсинаптической мембраны, взаимодействует с ее веществом. В результате этого взаимодействия структура постсинаптической мембраны временно изменяется, проницаемость ее для ионов натрия повышается, что приводит к перемещению ионов и, как следствие, появлению возбуждающего постсинаптического потенциала. Когда этот потенциал достигает определенной величины, возникает распространяющее возбуждение — потенциал действия.
 Рис. 29. Субмикроскопическое строение синапса:
Рис. 29. Субмикроскопическое строение синапса:
1 — пресинаптическая мембрана; 2 — постсинаптическая мембрана; 3 — синаптическая щель; 4 — синаптические пузырьки; 5 — нейропротофибриллы; 6 — митохондрии.
Через очень короткое время (несколько миллисекунд) медиаторы разрушаются специальными ферментами.
В настоящее время подавляющее большинство нейрофизиологов признает существование в спинном мозге и в различных отделах головного мозга двух качественно различных типов синапсов — возбуждающих и тормозящих. Полагают, что в специализированных тормозящих нейронах, в нервных окончаниях аксонов вырабатывается особый медиатор, оказывающий тормозящее воздействие на последующий нейрон. Природа этого медиатора еще окончательно не установлена. В коре больших полушарий таким медиатором некоторые авторы считают гамма-амино-масляную кислоту.
Под влиянием приходящего по аксону тормозящего нейрона импульса в синаптическую щель выделяется медиатор, который вызывает специфические изменения в постсинаптической мембране. Суть этих изменений состоит в том, что медиатор торможения, взаимодействуя с веществом постсинаптической мембраны, увеличивает ее проницаемость для ионов калия и хлора. Внутри клетки относительно увеличивается число анионов. В результате происходит не снижение величины внутреннего заряда мембраны (как это имеет место в возбуждающем синапсе), а повышение внутреннего заряда постсинаптической мембраны. Это ведет к возникновению тормозного постсинаптического потенциала, в результате чего наступает торможение.
В каждой нервной клетке расположено множество возбуждающих и тормозных синапсов. Это создает условия для их взаимодействия.
Статья на тему Физиология возбудимых тканей
25.Нарисуйте изменения мп на катоде при возникновении возбуждения, как изменяется возбудимость клетки во время ее возбуждения. Нарисуйте график изменения возбудимости при возбуждении клетки.
1.Что
такое возбуждение, какие ткани относятся
к возбудимым?
Возбуждение
– это процесс генерации потенциала
действия под воздействием пороговых и
сверхпороговых раздражителей. Возбудимые
ткани: мышечная, нервная и железистая.
2.Какую
функцию выполняет возбуждение. Приведите
примеры.
Возбуждение в
возбудимых тканях запускает специальные
реакции. Мышцы
– сокращение, нервы – импульс, выделение
медиатора, железа – секреция.
3.Какой
феномен отражает состояние возбуждения
мышечной клетки?
Заряд
клеточной мембраны
4.Что
такое возбудимость?
Способность
к возбуждению.
5.Как
можно оценить возбудимость различных
клеток, приведите пример?
По
параметру пороговой силы. Чем меньше
пороговая сила,тем больше возбудимость.
Самая
возбудимая ткань – нервная.
6.У
клетки А КУД 60 мв, мембранный потенциал
80 мв, у клетки В КУД 60 мв, мембранный
потенциал 90 мв, какая клетка является
более возбудимой, почему?
Клетка
А обладает большей возбудимостью, так
как возбуждение оценивает по пороговой
силе раздражителя(его минимальная сила,
при которой клетка возбуждается).
7.У
клетки А КУД 60 мв, мембранный потенциал(МП)
80 мв, у клетки В КУД 70 мв, МП 90 мв, какая
клетка является более возбудимой, ответ
докажите?
Клетки
обладают одинаковой возбудимостью, так
как возбуждение оценивает по пороговой
силе раздражителя(его минимальная сила,
при которой клетка возбуждается), а они
у клеток одинаковы.
8.Какие
электрофизиологические характеристики
клеточной мембраны предопределяют
возбудимость клеток? Приведите пример.
Мембранный
потенциал и КУД.
9.Приведите
пример реагирования возбудимой ткани
на пороговые и сверхпороговые раздражители
по закону «силовых соотношений».
Объясните причину такого реагирования.
Возбудимая
ткань на пороговые и сверхпороговые
раздражители реагирует по принципу чем
больше ток, тем больше ответная реакция.
Отдельные
клетки в силу разных причин обладают
разным порогом возбудимости, поэтому
в начале в мышце сокращаются наиболее
возбудимые клетки (пороговое сокращение),
а затем по мере увеличения силы
раздражителя в сокращение вовлекаются
все новые и новые менее возбудимые
клетки. Когда все клетки возбуждены,
увеличение амплитуды раздражителя не
вызывает прироста ответной реакции
(сокращения
10.Приведите
пример реагирования возбудимой ткани
по закону «все или ничего». Объясните
причину такого реагирования.
На
пороговые и сверхпороговые раздражители
ткань реагируют сокращением одинаковой
силы. Это характерно для однородных
систем (1 клетка, также сердечная.
Мышца сердца представляет собой так
называемый функциональный
синцитий,
а скелетная мышца – симпласт.
11.Что
отражает МПП нейрона, чему он равен, как
можно определить его величину.
Разность
потенциалов между наружной и внутренней
поверхностями мембраны клетки в покое.
12.Охарактеризуйте
ионные механизмы, обеспечивающие
возникновение мембранного потенциала
в нервных и мышечных клетках?
Калиевый
механизм – основной механизм, который
лежит в основе поляризации клеточной
мембраны. Также играет роль K
– Na
насос и ток Na
из клетки.
13.Как
изменится МПП при повышении внеклеточной
концентрации ионов К+, как это отразится
на возбудимости клетки?
МПП
снизится, так как снизится разность
зарядов. Возбудимость увеличится
вследствие изменения МПП.
14.Как
изменится МПД после обработки клетки
блокатором потенциалуправляемых
натриевых каналов мембраны?
снизится
15.Решите
задачу – внутриклеточный потенциал
мышечной клетки равен -80 мВ. Чему равен
мембранный потенциал покоя?
80
мВ.
16.Расшифруйте
понятия – поляризация, деполяризация,
реполяризация, гиперполяризация.
Поляризация
– ассиметричное распределение зарядов
относительно клеточной мембраны.
Деполяризация
– уменьшение напряжения на мембране.
Гиперполяризация
– увеличение напряжения на мембране.
Реполяризация
– восстановление мембранного потенциала
после де- и гиперполяризации.
17.Нарисуйте
электрограмму возникновения МПД при
пороговом и сверпорпоговом раздражении
мышечной клетки.
локальный ответ
(пороговая деполяризация)фаза быстрой
деполяризацииперескок (овершут)
– фаза перезарядкифаза реполяризации
следовой
отрицательный потенциалследовой
положительный потенциал
18.Объясните
ионный механизм возникновения МПД.
Идет
открытие управляемых натриевых каналов
под влиянием концентрационного и
электрического градинетов. Число
управляемых натриевых каналов больше
неуправляемых калиевых. В результате
происходит перезарядка мембраны. В фазу
перескока ток К из клетку начинает
преобладать над током Na
в клетку и заряд начинает падать.
19.Что
понимают под электротоном, какие
различают виды электротона?
Электотон
–
пассивные изменения мембранного
потенциала в области полюсов. Различают
анэлектротон, катэлектротон и
католическую депрессию.
20.Охарактеризуйте
динамику изменения возбудимости ткани
при ее длительной поляризации подпороговым
катодическим током.
Возбудимость
увеличивается, а проводимость снижается.
При продолжительном действии возникает
катодическая депрессия (снижается
возбудимость и проводимость).
21.Охарактеризуйте
динамику изменения возбудимости ткани
при ее длительной поляризации подпороговым
анодическим током.
Возбудимость
снижается, проводимость тоже. В результате
снижается функциональная деятельность.
Если действие тока растягивается, то
действие изменяется на строго
противоположное (анодическая депрессия):
повышается функциональная активность,
проводимость и возбудимость.
22.Нарисуйте
изменения мембранного потенциала
на катоде при кратковременном включении
подпорогового тока. Каковы механизмы
возникшей реакции.
МП
будет падать, так как введено большое
количество отрицательных зарядов.
23.Нарисуйте
изменения мембранного потенциала
на аноде при кратковременном включении
подпорогового тока. Каковы механизмы
возникшей реакции.
Мембранный
потенциал увеличивается.
24.Как
доказать полярный закон?
Полярный
закон: при прохождении через возбудимые
ткани постоянного ЭТ пороговой и
сверхпороговой величины при замыкании
цепи возникает под катодом, а при
размыкании – под анодом.
При
включении цепи мышца сокращается, т.е.
возбуждение проходит под каким-либо
электродом.
При
выключении цепи мышца сокращается, т.е.
возбуждение проходит под каким-то
электродом.
П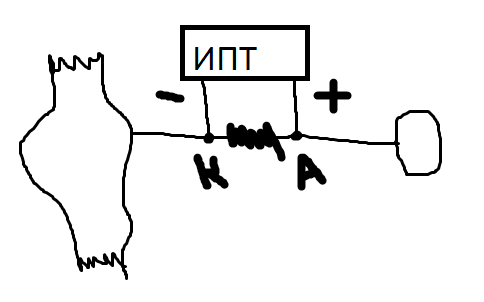 ри
ри
включении цепи мышца сокращается. При
выключении сокращения нет, т.к. возбуждение
не проходит блок.
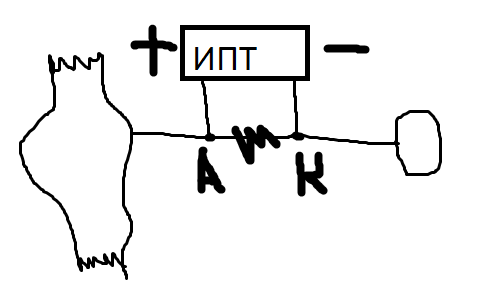
При
включении сокращения нет, так как оно
не проходит блок. при выключении
сокращение есть.
МП
снижается. Катод запускает деполяризацию,
после которой запускается процесс
регенерат. деполяризации.
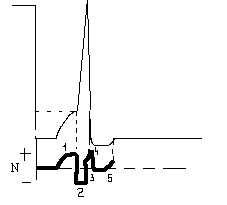
N
– нормальный уровень возбудимости
+повышение
возбудимости
— снижение
возбудимости
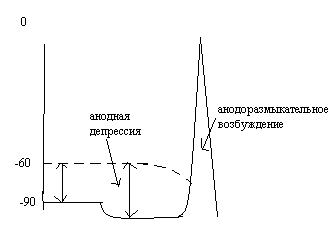
26.Нарисуйте изменения мп на аноде при возникновении возбуждения.
МП увеличивается,
при наступлении депрессии КУД тоже
снижается. При прекращении действия
тока наступает реполяризация и происходит
«перекрест» МП и КУД -> генерация ПД
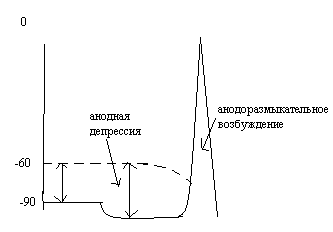
27.Объясните
механизм катодозамыкательного возбуждения
и анодоразмыкательного возбуждения.
См.вопросы
26 и 25
28.Охарактеризуйте динамику изменения возбудимости ткани при ее длительной поляризации плавнонарастающим по амплитуде катодическим током. Объясните причины возникающих изменений.
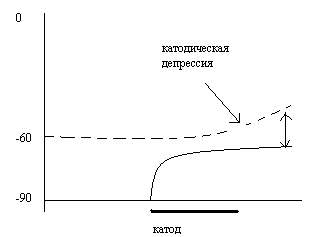
При
действии раздражителей, амплитуда
которых плавно нарастает, ткань
приспосабливается и может быть очень
большая амплитуда без наступления
возбуждения.
29.Охарактеризуйте
значение фактора времени действия
раздражителя на ткань для получении
возбуждения.
Чем длительно
раздражение, тем сильнее ответная
реакция возбудимой ткани. Для возникновения
возбуждение недостаточно действовать
на возбудимую ткань только силой
раздражения – необходимо, чтобы
раздражение длилось какое то время.
30.Как соотносятся между собой минимальная продолжительность действия тока на ткань и величина порогового значения тока?
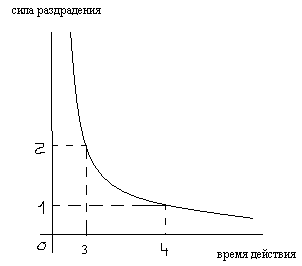
Чем больше сила
раздражения,тем меньше требуется времени
для возникновения возбуждения.
0-4 – полезное время
0-3 – хронаксия
31.Что такое полезное время, что такое хронаксия.
Наименьшая
длительность раздражения силой в одну
реобазу необходимая для возникновения
возбуждения называется полезным
временем.
Реобаза – та минимальная сила раздражение,
на которую при практически неограниченном
большой длительностью ее действия
возникнет минимальная ответная реакция.
Хронаксия
– минимальное время в течении которого
раздражитель с силой равной двум реобазам
вызывает возбуждение
32.На нерв действует раздражитель, который не влияет на куд, однако пороговый потенциал нервных волокон увеличился. Как это можно объяснить? Как изменится в этом случае возбудимость нерва?
Возбудимость
уменьшится, так как пороговый потенциал
увеличится.
Причина:
изменился заряд мембраны.
33.При нанесении раздражения на мышцу она не сокращается. О чем это свидетельствует? Дайте два варианта решения задачи.
1.Сила
раздражителя не достигла пороговой.
34.Охарактеризуйте типы сокращений скелетной мышцы в зависимости от частоты стимуляции двигательного нерва. Оптимум. Пессимум.
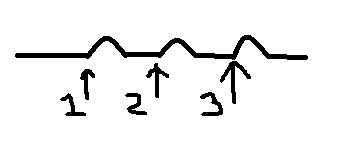
При
раздражении импульсами с малой частотой
наблюдаются одиночные
сокращения.
П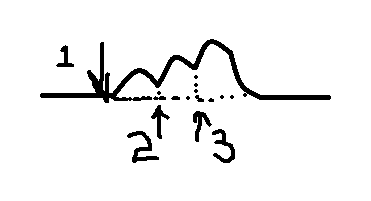 ри
ри
увеличении частоты при попадании каждого
последующего стимула на фазу расслабления
мышцы в предыдущем сокращении получается
зубчатый
тетанус (неполная
суммация).
При
еще большем увеличении получается
гладкий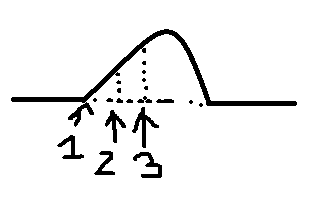
тетанус (полная
суммация).
Н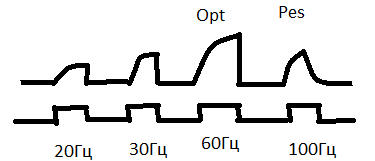 аиболее
аиболее
оптимальная частота раздражения мышц,
при которой она сокращается с наибольшей
амплитудой, называется оптимумом.
Если увеличивать
частоту раздражения дальше, то наступает
ухудшение сокращения мышцы или даже
полное его прекращение – пессимум
сокращения.
Оптимальное
сокращение зависит от концентрации Са
в зоне протофибрилл. Чем она больше, тем
лучше сокращение.
При пессимальном
сокращении стойкая деполяризация в
постсинаптической мембране смещается
на внесинаптическую мембрану, что ведет
к увеличению уровня КУД.
Возбудимость сердца относится к величине входящего тока, необходимого миоцитам или клеткам миокарда, клеткам в мышечном среднем слое сердца, для деполяризации или создания потенциала действия. Деполяризуется он или нет, зависит от того, возбудимы ли его потенциалзависимые «натриевые ионные каналы» или нет.
Более возбудимая клетка может иметь больше своих ионных каналов Na + в состоянии готовности, и даже если бы в нее поступал относительно слабый ток ионов Na +, клетка могла бы легко деполяризоваться.С другой стороны, в менее возбудимой клетке большая часть ионных каналов Na + может быть инактивирована, поскольку они не будут открываться в ответ на стимулы, представленные этим маленьким шариком, застрявшим в отверстии, и лишь некоторые из них готовы, и для этого потребуется сильный ток ионов Na +, прежде чем он будет деполяризован.
Допустим, это миоцит в одном из желудочков, а это график мембранного потенциала во времени. Во-первых, несколько положительных ионов, таких как натрий и кальций, проходят через щелевые контакты и входят в клетку, повышая мембранный потенциал до порогового уровня — обычно около 70 мВ.В этот момент открываются управляемые по напряжению каналы Na +, и множество ионов Na + устремляется в клетку, вызывая деполяризацию. Сразу после деполяризации, примерно при +20 мВ, каналы становятся неактивными, делая эти каналы недоступными для следующей деполяризации. После подъема вверх наступает плато, а затем, когда клетка реполяризуется, натриевые каналы начинают восстанавливаться, и даже если они закрыты, они все еще возбудимы, и в конечном итоге клетка реполяризуется обратно в обычное состояние около -90 мВ..
В течение большей части потенциала действия миокардиальная клетка не может снова деполяризоваться, и это называется периодом абсолютной рефрактерности. Другими словами, в течение периода абсолютной рефрактерности практически все натриевые каналы миоцита инактивируются, поэтому даже если пучок внутреннего тока исходит от соседней клетки, он буквально не может деполяризоваться. На каждой имеется много каналов Na +. миокардиальная клетка и каждый канал Na + работают независимо, но в целом большинство из них остаются инактивированными после восходящего движения через плато и до тех пор, пока клетка не реполяризуется примерно до -50 мВ, после чего некоторые каналы начинают восстанавливаться, после чего точка, клетка ответит на раздражитель..
Теперь способ измерения абсолютного рефрактерного периода состоит в том, что электрофизиолог подает ток на клетку миокарда, и в ответ она имеет потенциал действия. Пока этот потенциал действия проявляется, электрофизиолог подает в клетку несколько импульсов тока через равные промежутки времени. Если из-за всплеска тока ничего не происходит, это означает, что ячейка все еще находится в периоде абсолютной рефрактерности. Но если
.
Создать график онлайн и найти кратчайший путь или использовать другой алгоритм
Создать график и найти кратчайший путь. На странице справки вы найдете обучающее видео.
Установить матрицу смежности. Используйте запятую «,» в качестве разделителя
Матрица мультиграфа содержит вес минимальных ребер между вершинами.
Матрица неверна. Используйте запятую «,» в качестве разделителя. Матрица должна быть квадратной
Установите матрицу инцидентности.Используйте запятую «,» в качестве разделителя.
Матрица неверна. Используйте запятую «,» в качестве разделителя.
Ваш алгоритм отправлен на проверку и в случае успеха будет добавлен на сайт.
Невозможно создать график. Матрица смежности имеет неправильный формат. Нажмите кнопку «исправить матрицу», чтобы исправить матрицу, или кнопку «справка», чтобы открыть справку о формате матрицы смежности
Невозможно создать график. Матрица заболеваемости имеет неправильный формат. Нажмите кнопку «исправить матрицу», чтобы исправить матрицу, или кнопку «справка», чтобы открыть справку о формате матрицы заболеваемости
Выделяйте и перемещайте объекты мышью или перемещайте рабочее пространство.
Перетащите курсор для перемещения объектов
Выделяйте и перемещайте объекты мышью или перемещайте рабочее пространство.
Перетащите курсор для перемещения объектов
Щелкните в рабочей области, чтобы добавить новую вершину. Перечисление вершин
Выбрать первую вершину ребра
Выбрать вторую вершину ребра
Выбрать начальную вершину кратчайшего пути
Выбрать конечную вершину кратчайшего пути
Наименьшая длина пути% d
Путь не существует
Нажмите на объект, чтобы удалить
Добавить край
Направленный
Неориентированный
Матрица смежности
Сохранить
Отмена
наименьшее расстояние
Матрица заболеваемости
График сохранения
закрыть
Количество подключаемых компонентов
Количество слабосвязных компонент
Что вы думаете о сайте?
Имя (адрес электронной почты для обратной связи)
Обратная связь
Отправить
Чтобы задать нам вопрос или отправить комментарий, напишите нам по телефону
исправить матрицу
справка
Матрица имеет неправильный формат
Сохранить изображение графика
Полный отчет
Краткое сообщение
График не имеет эйлерова цикла
График имеет эйлеров цикл
Обработка…
Добавить вершину
Переименовать вершину
Переименовать
и
Изменить вес
не имеет веса
Переименовать группу
Голосовать
Рекомендовать алгоритмы
График не имеет эйлерова пути
График имеет эйлеров путь
График минимальных расстояний
Отметьте, чтобы сэкономить
Показать матрицу расстояний
Матрица расстояний
Выбрать источник максимального расхода
Выбрать мойку максимального расхода
Максимальный расход от% 2 до% 3 равен% 1
Поток из% 1 в% 2 не существует
Источник
Мойка
Граф не имеет гамильтонова цикла
График имеет гамильтонов цикл
График не имеет гамильтонова траектория
График имеет гамильтонов путь
Выбрать начальную вершину обхода
Порядок обхода:
Отвод края
Отменить
Сохранить график
По умолчанию
Vertex Style
Edge Style
Цвет фона
Multigraph поддерживает не все алгоритмы
не имеет веса
Используйте Cmd⌘ для выбора нескольких объектов.
Используйте Ctrl для выбора нескольких объектов.
Группа перетаскивания.
Группа копий
Удалить группу
Поиск в ширину
Раскраска графика
Найти подключенные компоненты
Поиск в глубину
Найдите цикл Эйлера
Найдите эйлеров путь
Алгоритм Флойда – Уоршалла
Упорядочить график
Найти гамильтонов цикл
Найти гамильтонов путь
Найти максимальный расход
Поиск минимального остовного дерева
Визуализация по весу
Радиус и диаметр поисковой диаграммы
Найдите кратчайший путь с помощью алгоритма Дейкстры
Вычислить степень вершин
.
Измерения центральности — документация по графическому инструменту 2.33
Вычислите PageRank каждой вершины.
- Параметры
- г
График Используемый график.
- демпфирование поплавок, опция (по умолчанию: 0,85)
Коэффициент демпфирования.
- чел.
VertexPropertyMap, необязательно (по умолчанию: нет) Вектор персонализации. Если опущено, постоянное значение \ (1 / N \)
будет использоваться.- вес
EdgePropertyMap, необязательно (по умолчанию: нет) Вес края. Если опущено, будет использоваться постоянное значение 1.
- prop
VertexPropertyMap, необязательно (по умолчанию: нет) Карта свойств вершин для хранения значений PageRank. В случае поставки он будет
использоваться без инициализации.- epsilon float, необязательно (по умолчанию: 1e-6)
Условие сходимости.Итерация остановится, если общая дельта всех
вершины ниже этого значения.- max_iter int, необязательно (по умолчанию: нет)
Если указано, это ограничит общее количество итераций.
- ret_iter bool, необязательный (по умолчанию: False)
Если true, также возвращается общее количество итераций.
- г
- Возвращает
- pagerank
VertexPropertyMap Карта свойств вершины, содержащая значения PageRank.{+} (u) = \ sum_ {y} A_ {u, y} w_ {u \ to y} \) переопределяется как сумма
весов выходящих из u ребер.Если у узла нет нулевой степени, предполагается, что он подключается ко всем остальным узлам.
с весом, пропорциональным \ (p (v) \), или постоянной, если нет
дается персонализация.Реализованный алгоритм последовательно повторяет вышеуказанные уравнения, пока
он больше не меняется согласно параметру epsilon. Оно имеет
время работы в зависимости от топологии.Если включено во время компиляции, этот алгоритм работает параллельно.
Список литературы
http://en.wikipedia.org/wiki/Pagerank
П. Лоуренс, Б. Сергей, М. Раджив, В. Терри,
«Рейтинг цитирования pagerank: наведение порядка в Интернете», Технические
отчет, Стэнфордский университет, 1998 г.- Langville-Survey-2005
А. Н. Лэнгвилл, К. Д. Мейер, «Обзор
Методы собственных векторов для поиска веб-информации », SIAM Review, vol. 47,
нет. 1, стр. 135-161, 2005, DOI: 10.1137 / S0036144503424786 [sci-hub, @tor]- adamic-polblogs (1,2)
л.А. Адамич, Н. Глэнс, «Политическая блогосфера.
и выборы в США 2004 г. », в материалах семинара WWW-2005 по вопросам
Экосистема веб-журналов (2005 г.). DOI: 10.1145 / 1134271.1134277 [sci-hub, @tor]
Примеры
>>> g = gt.collection.data ["polblogs"] >>> g = gt.GraphView (g, vfilt = gt.label_largest_component (g)) >>> pr = gt.pagerank (g) >>> gt.graph_draw (g, pos = g.vp ["pos"], vertex_fill_color = pr, ... vertex_size = gt.prop_to_size (пр, mi = 5, ma = 15), ... vorder = pr, vcmap = matplotlib.cm.gist_heat, ... output = "polblogs_pr.pdf") <...>
Теперь с вектором персонализации и весом краев:
>>> d = g.degree_property_map ("всего") >>> периферия = d.a <= 2 >>> p = g.new_vertex_property ("двойной") >>> p.a [периферия] = 100 >>> pr = gt.pagerank (g, pers = p) >>> gt.graph_draw (g, pos = g.vp ["pos"], vertex_fill_color = pr, ... vertex_size = gt.prop_to_size (pr, mi = 5, ma = 15), ... vorder = pr, vcmap = matplotlib.cm.gist_heat, ... output = "polblogs_pr_pers.pdf") <...>Персонализированные значения PageRank сети политических блогов
[adamic-polblogs], где даны вершины с очень низкой степенью
искусственно завышенные оценки..Подготовка к экзамену
IELTS — линейный график
- Дом
- Написание IELTS
- Линейный график
Линейные графики можно использовать, чтобы показать, как изменяются информация или данные
со временем. У них есть ось x (горизонтальная) и ось y (вертикальная). Обычно ось абсцисс
показывает период времени, а ось Y показывает, что измеряется. Линейные графики могут
использоваться при построении данных с пиками (подъемами) и минимумами (падениями). В
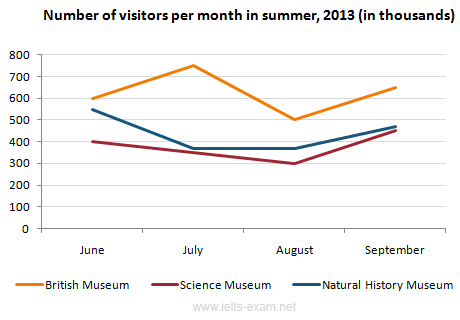
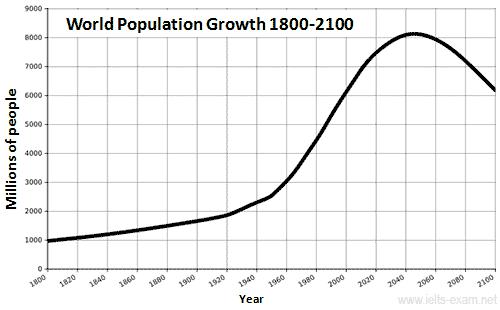
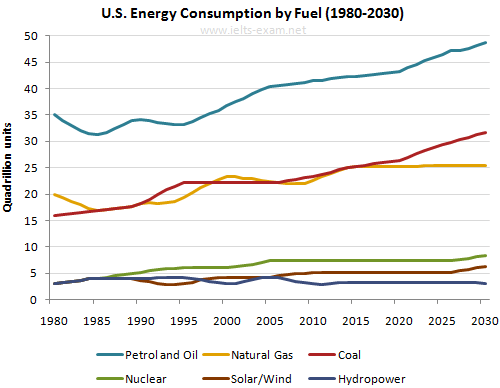
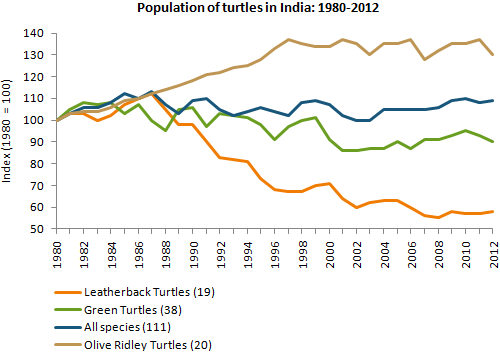
другими словами, они подчеркивают тенденции.Примеры вопросов

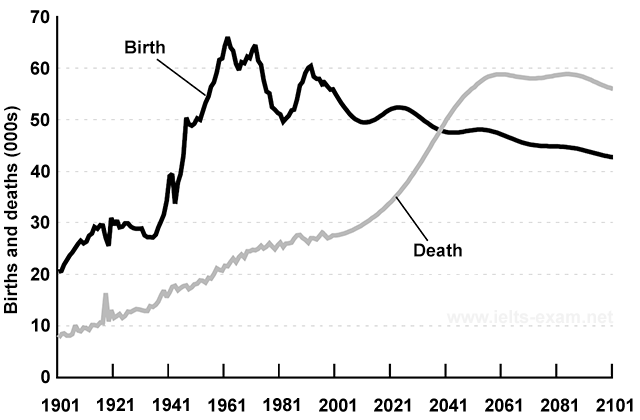
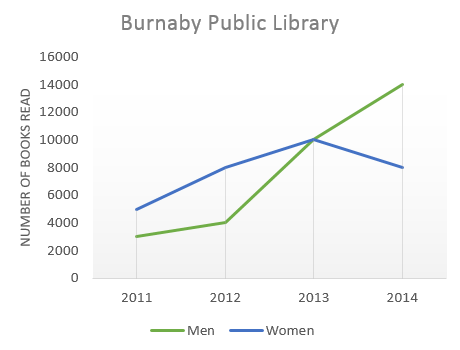
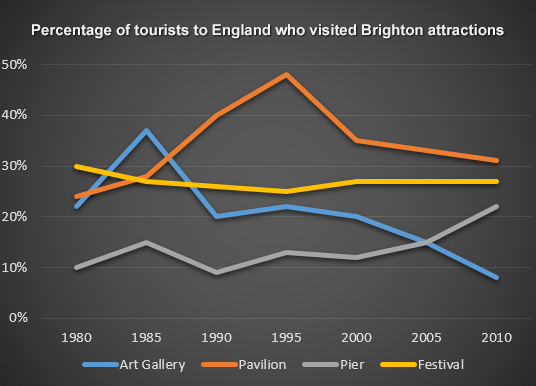
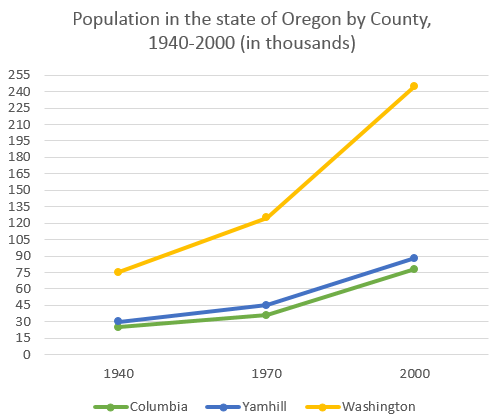
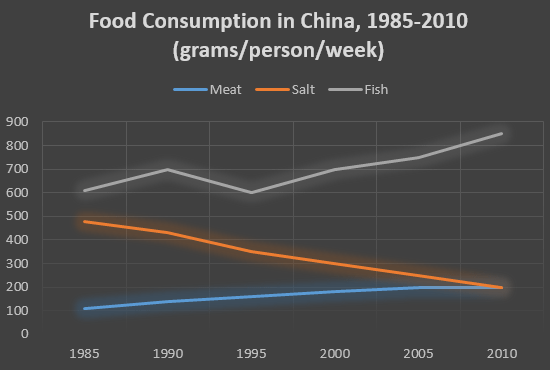
На графике представлена информация об изменениях показателей рождаемости и смертности в Новой Зеландии между 1901 и 2101 годами.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показано, как пожилые люди в США проводили свободное время с 1980 по 2010 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показаны численность населения Индии и Китая с 2000 года и прогнозируемый прирост населения до 2050 года.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показаны уровни загрязнения в Лондоне между 1600 и 2000 годами.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показаны относительные изменения цен на свежие фрукты и овощи, сахар и сладости, а также газированные напитки с 1978 по 2009 годы.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показано количество книг, прочитанных мужчинами и женщинами в публичной библиотеке Бернаби с 2011 по 2014 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
Линейный график показывает процент туристов в Англии, которые посетили четыре различные достопримечательности Брайтона.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показано количество денег, потраченных на покупку книг в Германии, Франции, Италии и Австрии в период с 1995 по 2005 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
График показывает изменение численности населения между 1940 и 2000 годами в трех разных округах США.Южный штат Орегон.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показаны изменения в потреблении продуктов питания китайцами в период с 1985 по 2010 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На линейном графике представлена информация о количестве посетителей трех лондонских музеев в период с июня по сентябрь 2013 г.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике показаны показатели переработки отходов в США с 1960 по 2011 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
Графики предоставляют информацию о численности населения мира и численности городского населения в различных регионах мира.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике представлена информация из Annual Energy Outlook 2008 о потреблении энергии в США с 1980 года с прогнозом до 2030 года.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
На графике представлена информация о прибытии международных туристов в пять стран.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
График показывает численность различных видов черепах в Индии в период с 1980 по 2012 год.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
Напишите отчет для преподавателя университета с описанием информации на графике.
Обобщите информацию, выбрав и сообщив об основных характеристиках, и при необходимости проведите сравнения.
Связанные темы
.
- pagerank