Глюкоза — Вікіпедія
| Глюкоза D-glucose | |
|---|---|
 | |
 | |
| Назва за IUPAC | (2R,3S,4R,5R)-2,3,4,5,6-Pentahydroxyhexanal |
| Інші назви | Виноградний цукор Blood sugar Dextrose Corn sugar D-Glucose Grape sugar |
| Ідентифікатори | |
| Абревіатури | Glc |
| Номер CAS | 50-99-7 |
| PubChem | 5793 |
| Номер EINECS | 200-075-1 |
| KEGG | C00031 |
| Назва MeSH | Glucose |
| ChEBI | 4167 |
| RTECS | LZ6600000 |
| Код ATC | V06DC01, B05CX01 і V04CA02 |
| SMILES | OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O |
| InChI | 1/C12h34O12/c7-1-2-3(8)4(9)5(10)6(11)12-2/h3-11H,1h3/t2-,3-,4+,5-,6+/m1/s1 |
| Номер Бельштейна | 1281604 |
| Номер Гмеліна | 83256 |
| 3DMet | B04623 |
| Властивості | |
| Молекулярна формула | C6H12O6 |
| Молярна маса | 180,16 г/моль |
| Молекулярна маса | 180,063388 |
| Густина | 1,54 г/см3 |
| Тпл | α-D-glucose: 146 °C β-D-glucose: 150 °C |
| Розчинність (вода) | 91 g/100 mL |
| Термохімія | |
| Ст. ентальпія утворення ΔfH | −1271 кДж/моль[1] |
| Ст. ентальпія згоряння ΔcH | −2805 кДж/моль |
| Ст. ентропія S | 209,2 Дж/(K·моль)[2] |
| Теплоємність, c | 218,6 Дж/(K·моль)[2] |
| Фармакологія | |
| Код ATC | B05 CX01 |
| Небезпеки | |
| MSDS | ICSC 0865 |
| Індекс ЄС | not listed |
| Якщо не зазначено інше, дані наведено для речовин у стандартному стані (за 25 °C, 100 кПа) | |
| Інструкція з використання шаблону | |
| Примітки картки | |
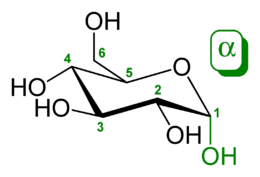
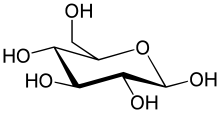
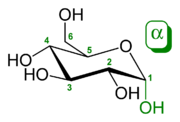
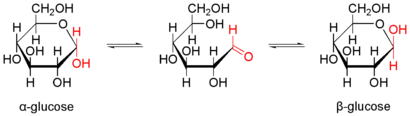 Перетворення між α- і β-D-Глюкозою відбувається через відкриту форму
Перетворення між α- і β-D-Глюкозою відбувається через відкриту форму
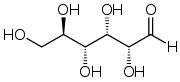
Глюко́за (від грец. γλυκύς — солодкий) (виноградний цукор, декстроза), С6Н12О6 — важливий моносахарид; безбарвні кристали солодкі на смак, легко розчиняються у воді.
Міститься в соку винограду, в багатьох фруктах, а також у крові тварин і людей.
М’язова робота виконується головним чином за рахунок енергії, яка виділяється при окисненні глюкози.
Глюкоза отримується при гідролізі полісахаридів крохмалю і целюлози (під дією ферментів або мінеральних кислот).
Погано розчиняється в органічних розчинниках. Неелектроліт.
Використовується як засіб посиленого харчування або як лікарська речовина, при обробці тканини. Також препарати на основі глюкози та сама глюкоза використовується у медицині при визначенні наявності та типу цукрового діабету в людини.
Природна кристалічна глюкоза (виноградний цукор) являє собою циклічну альфа-формулу. При розчиненні в воді вона переходить в ланцюгову, а через неї в бета-форму; при цьому установлюється динамічна рівновага між усіма формами.
Бета-форма також може бути виділена в кристалічному вигляді; у водному розчині вона утворює рівноважну систему з іншими формами.
Ланцюгова форма існує лише в розчинах, причому в дуже невеликій кількості, а в вільному вигляді не виділена.
Ізомерні форми сполук, які здатні переходити одна в одну називають таутомерними формами чи таутомерами. Явище таутомерії дуже розповсюджене серед органічних сполук.
Ймовірно, глюкоза відома людині з давніх часів, оскільки вона кристалізується з меду. Однак в чистому вигляді її виділили набагато пізніше: німецький хімік Андреас Маргграф отримав її в 1747 році з виноградного соку. Жозеф Луї Пруст в 1801 році вивів кристали α- D -глюкози з того ж виноградного соку. Завдяки цим експериментам за глюкозою закріпилася назва виноградного цукру.
Використання глюкози в якості підсолоджувача пов’язано з тим, що під час Наполеонівських воєн були заблоковані поставки тростинного цукру з Вест-Індії . Ще в 1000 році до н. е. в Китаї з крохмалю ( ферментацією рису) отримували штучний підсолоджувач — дисахарид мальтозу . До кінця XVIII ст. в Європі було відомо, що крохмаль можна обробити кислотою і отримати солодку речовину. Саме це дозволило К. С. Кірхгофу нагріванням картопляного крохмалю з сірчаною кислотою отримати солодкий сироп. В результаті оптимізації процесу він отримав сироп, який кристалізувався при настоюванні. Крім того, була зроблена спроба пресувати одержувану масу в твердий продукт і в такому вигляді продавати. Однак організувати виробництво не вдалося, оскільки Наполеон зазнав поразки і поставки цукру були відновлені.
Процес отримання глюкози досліджував французький хімік Соссюр . Він з’ясував, що крохмаль піддається гідролізу , при якому руйнуються зв’язки між вуглеводними фрагментами, причому на кожен розрив витрачається по одній молекулі води. Також було виявлено, що цукерки, вироблені з сиропів глюкози, не такі солодкі, як ті, що отримують з сахарози. Тому в Німеччині та інших країнах Європи було організовано виробництво глюкози.
У той час промислова глюкоза була недостатньо чистою і виходила у вигляді сиропів. Її доводилося багато разів кристалізувати з води, або використовувати органічні розчинники. Єдиною твердою формою, яку випускали у великій кількості, була лита глюкоза: сироп заливали в форми, де він затвердівав.
У 1923 році в США Вільям Б. Ньюкірк ( англ. William B. Newkirk ) запатентував промисловий спосіб отримання глюкози. Цей спосіб відрізнявся ретельним контролем умов кристалізації, завдяки чому глюкоза випадала з розчину у вигляді чистих, великих кристалів.
Глюкоза може відновлютися йодоводнем до 2-йодгексану[3]:
HO−Ch3−(CH(OH))4−CH=O→HICh4−CHI−(Ch3)3−Ch4{\displaystyle {\ce {HO-Ch3-(CH(OH))4-CH=O ->[HI]Ch4-CHI-(Ch3)3-Ch4}}}
α-глюкоза взаємодіє з метанолом, утворюючи O-метил-α-глюкозу[3]. При цьому метильна груп приєднується до атома оксигену з тієї гідроксильної групи, яка у ланцюговій формі є альдегідною.
При окисненні утворюється глюкуронова кислота. Вона також може бути і в циклічній, і в ланцюговій формі.
Відновлює гідроксид міді(||) до оксиду міді(|), а реактив Толленса до аміаку та срібла.
При взаємодії з гідроксиламіном утворює оксим, який під дією оцтового ангідриду перетворюється на ацильований гідроксинітрил. Останній під дією оксиду срібла та аміаку перетворюється на арабінозу:
HO−Ch3−(CH(OH))4−CH=O+Nh3−OH→−h3OHO−Ch3−(CH(OH))4−CH=B−OH{\displaystyle {\ce {HO-Ch3-(CH(OH))4-CH=O +Nh3-OH ->[-h3O]HO-Ch3-(CH(OH))4-CH=B-OH}}}
2HO−Ch3−(CH(OH))4−CH=N−OH+5(Ch4−CO)2=O→−7h3O2h4C−CO−O−Ch3−(CH(O−CO−Ch4))4−C≡N{\displaystyle {\ce {2HO-Ch3-(CH(OH))4-CH=N-OH + 5(Ch4-CO)2=O->[-7H_2O]2h4C-CO-O-Ch3-(CH(O-CO-Ch4))4-C#N}}}
h4C−CO−O−Ch3−(CH(O−CO−Ch4))4−C≡N→Nh4Ag2OHO−Ch3−(CH(OH))3−CH=O{\displaystyle {\ce {h4C-CO-O-Ch3-(CH(O-CO-Ch4))4-C#N->[{Ag2O}][{Nh4}]HO-Ch3-(CH(OH))3-CH=O}}}
- ↑ Ponomarev, V. V.; Migarskaya, L. B. (1960). Heats of combustion of some amino-acids. Russ. J. Phys. Chem. (Engl. Transl.) 34: 1182–83.
- ↑ а б Boerio-Goates, Juliana (1991). Heat-capacity measurements and thermodynamic functions of crystalline α-D-glucose at temperatures from 10K to 340K. J. Chem. Thermodynam. 23 (5): 403–9. doi:10.1016/S0021-9614(05)80128-4.
- ↑ а б Ластухін, Воронов (2006). Органічна хімія. ISBN 966-7022-19-6.
| В іншому мовному розділі є повніша стаття Glucose (англ.). Ви можете допомогти проєкту, розширивши поточну статтю за допомогою перекладу з англійської. Натисніть [розгорнути] праворуч для перегляду інструкцій.
|
Гліколіз — Вікіпедія
Загальна схема гліколізу
Гліко́ліз або шлях Ембдена—Меєргофа—Парнаса (від дав.-гр. γλυκός — солодкий та λύσης — розщеплення) — ланцюг із десяти реакцій, внаслідок яких глюкоза C6H12O6 перетворюється на піруват, C3H3O-3 з утворенням АТФ (аденозинтрифосфату) та НАДН (відновленого нікотинамідаденіндинуклеотиду).
У аеробних організмів гліколіз йде перед циклом трикарбонових кислот та ланцюгом переносу електронів. Вони разом вивільняють більшу частину енергії, яка міститься в глюкозі. За аеробних умов піруват потрапляє в мітохондрії, де повністю окиснюється до СО2 та Н2О.
За недостатності кисню (це буває в м’язах, які активно скорочуються) піруват перетворюється на лактат. У деяких анаеробних організмах, наприклад, дріжджів, піруват перетворюється не на лактат, а на етанол. Утворення етанолу та лактату з глюкози — це приклади бродіння.



Дослідники гліколізу
Серед метаболічних шляхів гліколіз відкрили першим і він найдослідженіший . 1897 року брати Ганс[en] та Едуард Бюхнери виготовляли безклітинні екстракти дріжджів для лікування. Очевидно, вони при цьому не могли скористатися отруйними для людини антисептиками (наприклад, фенолом), тому спробували поширений у кулінарії консервант — сахарозу. З’ясувалося, що в дріжджових екстрактах ця речовина швидко зброджувалась до етилового спирту. Так було вперше встановлено, що бродіння може відбуватись поза межами живих клітин. 1907 року за це відкриття Едуарда Бюхнера відзначили Нобелівською премією з хімії[4].
Вивчення позаклітинного бродіння лишалося одним із основних завдань біохімії аж до 1940-х років. Перебіг цього метаболічного шляху в клітинах дріжджів описали Отто Варбург, Ганс фон Ейлер-Хельпін та Артур Гарден (два останні отримали Нобелівську премію з хімії 1929 року[5]). Гліколіз у м’язах дослідили Густав Ембден[en] та Отто Меєргоф (Нобелівська премія з медицини та фізіології 1922 року[6]). Свій внесок у дослідження гліколізу зробили Карл Нойберг[en], Яків Парнас, Герті та Карл Корі.
Завдяки вивченню гліколізу було зроблено «побічні» відкриття: винайшли численні методи очищення ферментів, з’ясували центральну роль АТФ та інших фосфорильованих сполук у метаболізмі, відкрили коензими, такі як НАД.
Поширення й значення[ред. | ред. код]
Гліколіз є майже універсальним шляхом катаболізму глюкози для всіх клітинних організмів. Ймовірно, він виник на ранніх етапах розвитку життя на Землі, ще до того, як в атмосфері накопичився кисень внаслідок діяльності фотосинтезуючих істот. В еволюційному плані послідовність гліколітичних реакцій — надзвичайно давній шлях. Його ферменти у хребетних тварин, дріжджів та рослин дуже споріднені як за амінокислотними послідовностями, так і за тривимірною структурою. Секвенування ДНК з геномів численних організмів показало, що тільки в деяких архей та паразитичних бактерій відсутні один або кілька ферментів гліколізу, попри те сам шлях зберігається, імовірно, у модифікованій формі.
В евкаріотичних клітинах реакції гліколізу відбуваються у цитозолі. У більшості таких клітин саме цей метаболічний шлях з-поміж інших посідає перше місце за кількістю атомів карбону, що зазнають у ньому перетворень.
У більшості аеробних евкаріотичних організмів гліколіз створює субстрат до подальшого окиснення в мітохондріях, де за наявності кисню запасається значна кількість енергії у вигляді АТФ. Втім для деяких типів клітин, які не мають мітохонондрій або перебувають далеко від капілярів, зокрема в еритроцитах ссавців та клітинах кришталика, гліколіз є головним джерелом енергії. Для скелетних м’язів та інших тканин в умовах дуже великого навантаження або гіпоксії гліколіз вигідний не лише тому, що дає можливість одержати енергію за умов нестачі кисню, а ще й тому, що відбувається дуже швидко і забезпечує синтез АТФ у 10,5 разів швидше, ніж аеробне окиснення органічних речовин[8]. Серед м’язових волокон виділяють групу волокон IIB, які містять мало мітохондрій, оточені невеликою кількістю капілярів та здатні до сильних швидких скорочень протягом невеликого часу. Такі волокна отримують більшу частину енергії за рахунок гліколізу. Клітини мозкового шару нирок, лімфоцити, клітини шкіри також є суттєво залежними від гліколізу.[9]
Для головного мозку ссавців (за винятком випадків голодування) характерне активне використання гліколізу навіть за наявності кисню під час ембріонального нейрогенезу[en], а також при рості дендритів та формуванні синаптичних контактів у дорослому мозку[11]. Гліколіз також є важливим джерелом енергії для сперматозоїдів. Сперматозоїди різних видів ссавців мають різну ефективність гліколізу: сперматозоїди людини здатні виживати та рухатися в анаеробному середовищі, тоді як клітини мурчака та свині знерухомлюються й без кисню гинуть; сперматозоїди биків та макак резусів займають проміжне положення.[12]
У рослин від гліколізу залежать, головним чином, тканини, спеціалізовані на зберіганні крохмалю (наприклад, бульби картоплі) та водяні рослини (настурція лікарська).
Іншими шляхами окиснення глюкози є пентозофосфатний шлях та шлях Ентнера — Дудорова. Останній є заміною гліколізу в деяких грам-негативних та, дуже рідко, грам-позитивних бактерій і має багато спільних із ним ферментів.
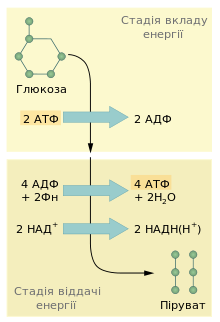 Узагальнена схема двох стадій гліколізу
Узагальнена схема двох стадій гліколізу
Традиційно гліколіз поділяють на дві стадії: підготовчу, яка вимагає поглинання енергії (п’ять перших реакцій), та стадію віддавання енергії (п’ять останніх реакцій). Інколи четверту та п’яту реакції виділяють в окрему проміжну стадію.
На першій стадії відбувається фосфорилювання глюкози у шостому положенні, ізомеризація одержаного глюкозо-6-фосфату до фруктозо-6-фосфату та повторне фосфорилювання вже у першому положенні. Внаслідок цього утворюється фруктозо-1,6-бісфосфат. Фосфатні групи на моносахариди переносяться з АТФ. Це необхідно для активації молекул — збільшення вмісту в них вільної енергії. Потім фруктозо-1,6-бісфосфат розщеплюється до двох фосфотріоз, які можуть вільно перетворюватись одна на одну.
На другій стадії (віддавання енергії) фосфотріоза (гліцеральдегід-3-фосфат) окиснюється та фосфорилюється неорганічним фосфатом. Утворюється продукт, що в серії екзергонічних реакцій, спряжених із синтезом чотирьох молекул АТФ, перетворюється на піруват. Отже, під час гліколізу відбувається три принципові перетворення:
Перша стадія[ред. | ред. код]
Фосфорилювання глюкози[ред. | ред. код]
Перша реакція гліколізу — фосфорилювання глюкози з утворенням глюкозо-6-фосфату, що каталізується ферментом гексокіназою. Донором фосфатної групи є молекула АТФ. Реакція відбувається тільки за наявності іонів Mg2+, тому що справжнім субстратом для гексокінази є не АТФ4-, а комплекс MgАТФ2-. Магній екранує негативний заряд фосфатної групи, таким чином полегшуючи здійснення нуклеофільної атаки на останній атом фосфору гідроксильною групою глюкози.

- ΔG0 = −16,7 кДж/моль
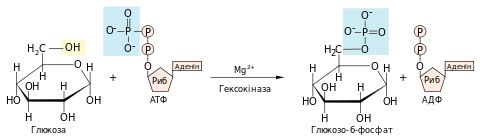
Внаслідок фосфорилювання відбувається не лише активація молекули глюкози, а й її «ув’язнення» всередині клітини: плазматична мембрана має білки-переносники для глюкози, але не для її фосфорильованої форми. Тому велика заряджена молекула глюкозо-6-фосфату не може пройти крізь мембрану, хоча її концентрація у цитоплазмі більша, аніж у позаклітинній рідині.
Фермент гексокіназа є майже у всіх організмів, основним його субстратом є глюкоза. Проте він може каталізувати фосфорилювання інших гексоз: D-фруктози, D-манози тощо. У людини є чотири ізоформи гексокінази (від I до IV). Один з ізоферментів — гексокіназа IV або глюкокіназа — відрізняється від інших форм особливостями кінетики та регулюванням своєї активності.
Ізомеризація глюкозо-6-фосфату[ред. | ред. код]
У другій реакції гліколізу відбувається ізомеризація глюкозо-6-фосфату до фруктозо-6-фосфату під впливом ферменту глюкозофосфатізомерази (гексозофосфатізомерази). Спочатку відбувається відкриття шестичленного піранозного кільця глюкозо-6-фосфату, тобто перехід цієї речовини у лінійну форму, після чого карбонільна група з першого положення переноситься в друге через проміжну ендіольну форму. Тобто альдоза перетворюється на кетозу. Утворена лінійна молекула фруктозо-6-фосфату замикається у п’ятичленне фуранозне кільце.
- ΔG0 = 1,7 кДж/моль
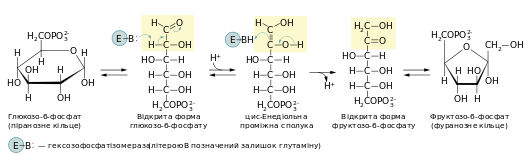
Через незначну зміну вільної енергії реакція є оборотною. Ізомеризація глюкозо-6-фосфату є необхідною умовою для подальшого гліколізу, оскільки наступна реакція — ще одне фосфорилювання — потребує гідроксильної групи в першому положенні.
Фосфорилювання фруктозо-6-фосфату[ред. | ред. код]
Після стадії ізомеризації починається друга реакція фосфорилювання, за якої фруктозо-6-фосфат перетворюється на фруктозо-1,6-бісфосфат[en] завдяки приєднанню фосфатної групи АТФ. Реакцію каталізує фермент фосфофруктокіназа-1[en] (скорочено ФФК-1). Існує також фермент ФФК-2[en], який каталізує утворення фруктозо-2,6-бісфосфату[en] іншим метаболічним шляхом .
- ΔG0 = −14,2 кДж/моль

У цитоплазмі ця реакція є незворотною. Вона першою вірогідно визначає розщеплення речовин по гліколітичному шляху, оскільки глюкозо-6-фосфат та фруктозо-6-фосфат можуть вступати в інші метаболічні перетворення, а фруктозо-1,6-бісфосфат використовується лише для гліколізу. Саме утворення фруктозо-1,6-бісфосфату є визначальною стадією гліколізу.
У рослин, деяких бактерій та найпростіших є також форма фосфофруктокінази, що використовує як донор фосфатної групи пірофосфат, а не АТФ. ФФК-1 як алостеричний фермент[en] зазнає складного регулювання. До позитивних модуляторів належать продукти розщеплення АТФ — АДФ та АМФ, рибулозо-5-фосфат[en] (проміжний продукт пентозофосфатного шляху), у деяких організмів — фруктозо-2,6-бісфосфат. Негативним модулятором є АТФ.
Розщеплення фруктозо-1,6-бісфосфату на дві фосфотріози[ред. | ред. код]
Фруктозо-1,6-бісфосфат розщеплюється до двох фосфотріоз: гліцеральдегід-3-фосфат та дигідроксиацетонфосфат під впливом фруктозо-1,6-фосфатальдолази (частіше просто альдолаза). Назва ферменту альдолази походить від зворотної реакції альдольної конденсації. Механізм проходження реакції зображений на схемі:
- ΔG0 = 23,8 кДж/моль
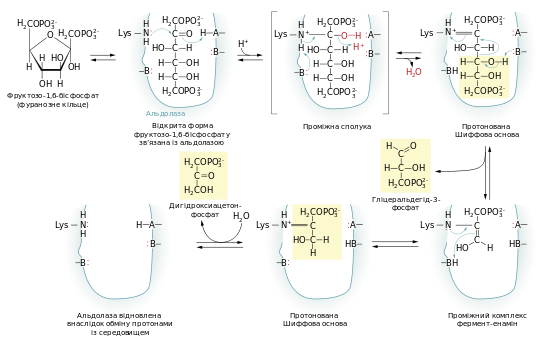
Хоча стандартна зміна вільної енергії під час розщеплення фруктозо-1,6-бісфосфату є позитивною й має велике абсолютне значення, в реальній клітині через низьку концентрацію фосфотріоз реакція легко проходить в обидва боки.
Описаний механізм реакції характерний тільки для альдолази класу I, поширеної у рослин та тварин. У клітинах бактерій та грибів є альдолаза класу II. Вона каталізує реакцію іншим шляхом.
Механізм реакції альдольного розщеплення демонструє важливість ізомеризації і в другій реакції гліколізу. Якби такому перетворенню підлягала альдоза (глюкоза), то утворилася б одна двокарбонова та одна чотирикарбонова сполука. Кожна з них мала б метаболізуватися своїм власним шляхом. Натомість трикарбонові сполуки, утворені при розщепленні кетози (фруктози), можуть легко перетворюватись одна на одну і метаболізуються один шляхом.
Ізомеризація фосфотріоз[ред. | ред. код]
У подальших реакціях гліколізу бере участь тільки одна із фосфотріоз, утворених з фруктозо-1,6-бісфосфату, а саме гліцеральдегід-3-фосфат. Проте інший продукт — дигідроксиацетонфосфат — швидко і зворотно може перетворюватись на гліцеральдегід-3-фосфат (каталізує цю реакція тріозофосфатізомераза).
- ΔG0 = 7,5 кДж/моль

Механізм реакції схожий на ізомеризацію глюкозо-6-фосфату у фруктозо-6-фосфат. Рівновага реакції зміщена в бік утворення дигідроксиацетонфосфату (96 %), проте через постійне витрачання гліцеральдегід-3-фосфату весь час відбувається зворотне перетворення.
Після перетворення двох «половинок» глюкози на гліцеральдегід-3-фосфат атоми карбону, що походять від її C-1, C-2 та C-3, стають хімічно тотожними C-6, C-5 та C-4 відповідно. Ця реакція завершує підготовчу стадію гліколізу.
Друга стадія[ред. | ред. код]
Окиснення гліцеральдегід-3-фосфату[ред. | ред. код]
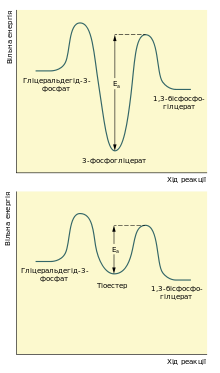 Зміна вільної енергії під час окиснення гліцеральдегід-3-фосфату та фосфорлиювання утвореного 3-фосфогліцерату, якщо вони відбуваються послідовно (зверху) та якщо вони спряжені завдяки ковалентному зв’язуванню проміжного продукту із ферментом (знизу).
Зміна вільної енергії під час окиснення гліцеральдегід-3-фосфату та фосфорлиювання утвореного 3-фосфогліцерату, якщо вони відбуваються послідовно (зверху) та якщо вони спряжені завдяки ковалентному зв’язуванню проміжного продукту із ферментом (знизу).
Першою реакцією стадії віддачі енергії гліколізу є окиснення гліцеральдегід-3-фосфату із одночасним його фосфорилюванням. Це здійснює фермент гліцеральдегід-3-фосфатдегідрогеназа. Альдегід перетворюється не на вільну кислоту, а на змішаний ангідрид із фосфатною кислотою (1,3-бісфосфогліцерат). Сполуки такого типу — ацилфосфати — мають дуже велику від’ємну зміну вільної енергії гідролізу (ΔG0 = −49,3 кДж/моль).
Реакцію перетворення гліцеральдегід-3-фосфату до 1,3-бісфосфогліцерату можна розглядати як два окремі процеси: окиснення альдегідної групи НАД+ та приєднання фосфатної групи до утвореної карбонової кислоти. Перша реакція термодинамічно вигідна (ΔG0 = −50 кДж/моль), друга, навпаки, невигідна.
Зміна вільної енергії для другої реакції майже така сама, тільки ця зміна додатна. Якби вони відбувались послідовно одна за одною, то друга реакція вимагала б надто великої енергії активації, щоб перебігати в клітині з достатньою швидкістю. Але обидва процеси є спряженими тому, що проміжна сполука — 3-фосфогліцерат — ковалентно зв’язана із залишком цистеїну тіоестерним зв’язком в активному центрі ферменту. Такий тип зв’язку дозволяє «законсервувати» частину енергії, яка виділяється під час окиснення гліцеральдегід-3-фосфату, та використати її для реакції із ортофосфатною кислотою.
- ΔG0 = 6,3 кДж/моль
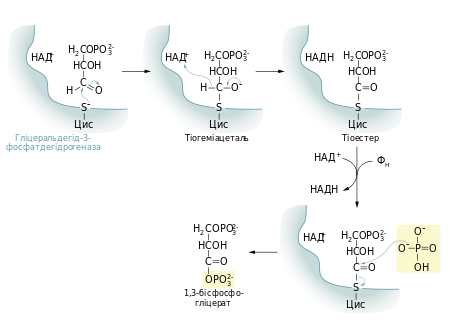
Для цієї стадії гліколізу необхідний кофермент НАД+. Його концентрація в клітині (менше 10−5М) значно менша, ніж кількість глюкози, що метаболізується за декілька хвилин. Тому в клітині постійно відбувається повторне окиснення НАД+.
Перенесення фосфатної групи 1,3-бісфосфогліцерату на АДФ[ред. | ред. код]
У наступній реакції великий запас енергії ацилфосфату використовується для синтезу АТФ. Ферменти фосфогліцераткіназа 1-го та 2-го типу (назва від зворотної реакції) каталізують перенесення фосфатної групи із 1,3-бісфосфогліцерату на АДФ з утворенням АТФ та 3-фосфогліцерату.
- ΔG0 = −18,6 кДж/моль

Такий тип синтезу АТФ, за якого використовується розчинна сполука з високим потенціалом перенесення фосфатної групи, називають сусбстратним фосфорилюванням на противагу окисному фосфорилюванню. Останнє має місце під час аеробного окиснення у внутрішній мембрані мітохондрій.
Шоста та сьома реакції гліколізу спряжені між собою і 1,3-бісфосфогліцерат є спільним проміжним продуктом. Перша з них сама по собі була б ендергонічною, проте витрати енергії компенсуються другою реакцією — виражено екзергонічною. Сумарне рівняння цих двох процесів можна записати так:
- Гліцеральдегід-3-фосфат + АДФ + Фн + НАД+ → 3-фосфогліцерат + АТФ + НАДH(H+), ΔG0 = −12,2 кДж/моль;
Слід зауважити, що для однієї молекули глюкози ця реакція відбувається двічі, оскільки з однієї молекули глюкози були утворені дві молекули гліцеральдегід-3-фосфату. Отже, на цьому етапі синтезуються дві молекули АТФ, і це компенсує енергетичні затрати першої стадії гліколізу.
Ізомеризація 3-фосфогліцерату[ред. | ред. код]
У восьмій реакції гліколізу фермент фосфогліцератмутаза у присутності іонів магнію каталізує перенесення фосфатної групи 3-фосфогліцерату з третього положення в друге. Внаслідок цього утворюється 2-фосфогліцерат. Реакція відбувається у два етапи. На першому з них фосфатна група, початково приєднана до залишку гістидину в активному центрі ферменту, переноситься на C-2 3-фосфогліцерату і утворюється 2,3-бісфосфогліцерат. На другому етапі фосфатна група у третьому положенні синтезованої сполуки переноситься на гістидин. Таким чином регенеруються фосфорильований фермент та продукується 2-фосфогліцерат.
- ΔG0 = 4,4 кДж/моль

Початкове фосфорилювання фосфогліцератмутази здійснюється реакцією із 2,3-бісфосфогліцерату, незначної концентрації якого достатньо для активації ферменту.
Дегідратація 2-фосфогліцерату[ред. | ред. код]
Наступна реакція — утворення енолу в результаті дегідратації (відщеплення води) 2-фосфогліцерату — веде до утворення фосфоенолпірувату (скорочено ФЕП). Процес каталізує фермент енолаза[en].
- ΔG0 = 7,5 кДж/моль
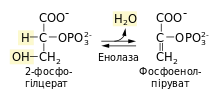
Це друга реакція утворення речовини із високим потенціалом перенесення фосфатної групи в процесі гліколізу. Зміна вільної енергії при гідролізі фосфатного естеру звичайного спирту значно нижча порівняно з такою зміною при гідролізі енолфосфату, зокрема для 2-фосфогліцерату ΔG0 = −17,6 кДж/моль, а для фосфоенолпірувату ΔG0 = −61,9 кДж/моль.
Перенесення фосфатної групи із ФЕП на АДФ[ред. | ред. код]
Останню реакція гліколізу — перенесення фосфатної групи з фосфоенолпірувату на АДФ — каталізує піруваткіназа[en] за наявності іонів K+ та Mg2+ або Mn2+. Продуктом цієї реакції є піруват, який спочатку утворюється в енольній формі, після чого швидко й неферментативно таутомеризується в кетонну форму.
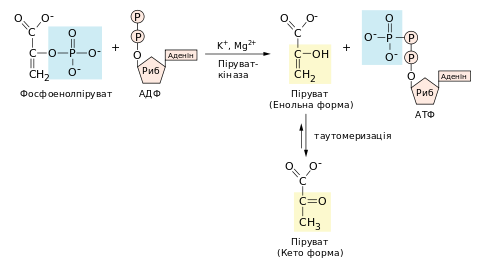
Реакція має велику від’ємну зміну вільної енергії, головним чином, завдяки екзергонічому процесу таутомеризації. Близько половини енергії (30,5 кДж/моль), що виділяється при гідролізі ФЕП (61,9 кДж/моль), використовується на субстратне фосфорилювання. Решта (31,5 кДж/моль) слугує рушійною силою, що штовхає реакцію в бік утворення пірувату та АТФ. За клітинних умов реакція є незворотною .
Сумарний вихід гліколізу[ред. | ред. код]
| Зміна вільної енергії в реакціях гліколізу в еритроцитах | ||
|---|---|---|
| Реакція | ΔG0 (кДж/моль) | ΔG (кДж/моль) |
| Глюкоза + АТФ → глюкозо-6-фосфат + АДФ | −16,7 | −33,4 |
| Глюкозо-6-фосфат ↔ фруктозо-6-фосфат | 1,7 | від 0 до 25 |
| Фруктозо-6-фосфат + АТФ → фруктозо-1,6-бісфосфат + АДФ | −14,2 | −22,2 |
| Фруктозо-1,6-бісфосфат ↔ гліцеральдегід-3-фосфат + дигідроксиацетонфосфат | 28,3 | від −6 до 0 |
| Дигідроксиацетонфосфат ↔ гліцеральдегід-3-фосфат | 7,5 | від 0 до 4 |
| Гліцеральдегід-3-фосфат + Фн + НАД+ ↔ 1,3-бісфосфогліцерат + НАДH + H+ | 6,3 | від −2 до 2 |
| 1,3-Бісфосфогліцерат + АДФ ↔ 3-фосфогліцерат + АТФ | −18,8 | від 0 до 2 |
| 3-Фосфогліцерат ↔ 2-фосфогліцерат | 4,4 | від 0 до 0,8 |
| 2-Фосфогліцерат ↔ фосфоенолпіруват + H2O | 7,5 | від 0 до 3,3 |
| Фосфоенолпіруват + АДФ → піруват + АТФ | −31,4 | −16,7 |
| Жовтим виділені реакції, незворотні в реальних умовах клітини | ||
Загальне рівняння гліколізу має такий вигляд:
- Глюкоза + 2Фн + 2АДФ + 2НАД+ → 2 піруват + 2АТФ + 2НАДН + 2Н+ + 2Н2О.
Сумарна кількість енергії, що виділяється внаслідок розщеплення глюкози до пірувату становить 146 кДж/моль, на синтез двох молекул АТФ витрачається 61 кДж/моль, решта (85 кДж/моль) перетворюється на тепло.
При повному окисненні глюкози до вуглекислого газу та води виділяється 2 840 кДж/моль. Якщо порівняти це із загальним виходом екзергонічних реакцій гліколізу (146 кДж/моль), стає зрозуміло, що 95 % енергії глюкози залишається «ув’язненою» у молекулах пірувату. Хоча реакції гліколізу є універсальними майже для всіх організмів, подальша доля його продуктів — пірувату та НАДН — відрізняється у різних живих істот та залежить від умов.
В аеробних організмів за достатньої концентрації кисню НАД+ регенерується шляхом передачі електронів в електронтранспортний ланцюг, який в еукаріот розташований у внутрішній мембрані мітохондрій. Кінцевим акцептором електронів при цьому є кисень. Піруват зазнає окисного декарбоксилювання, перетворюється у ацетил-КоА і надходить у цикл Кребса, де відбувається його подальше окиснення. Відщеплені електрони також потрапляють в електронтранспортний ланцюг.
З іншого боку в анаеробних умовах відновлений НАДН не може передати свої електрони на кисень. Тому є два варіанти їхнього перенесення:
Анаеробний метаболізм глюкози дає значно менше енергії, ніж аеробний.
Включення інших вуглеводів у процес гліколізу[ред. | ред. код]
Окрім глюкози у процесі гліколізу зазнають перетворень ще чимало вуглеводів. Найважливішими з них є полісахариди крохмаль та глікоген, дисахариди сахароза, лактоза, мальтоза та трегалоза, а також моносахариди — фруктоза, галактоза та маноза.
Полісахариди[ред. | ред. код]
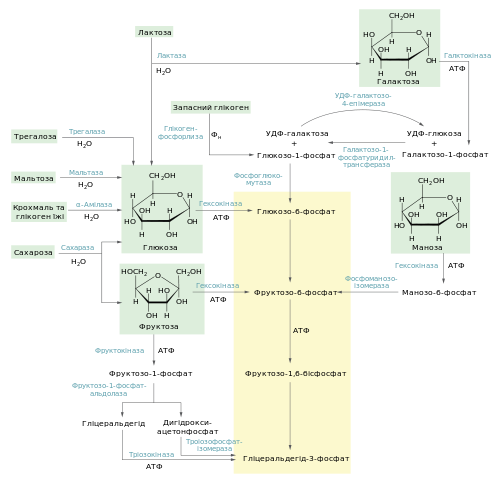 Схема включення у підготовчу фазу гліколізу глікогену, крохмалю, дисахаридів та гексоз
Схема включення у підготовчу фазу гліколізу глікогену, крохмалю, дисахаридів та гексоз
Полісахариди, що включаються в процес гліколізу, можуть мати різне походження, від чого залежить їхня доля. Крохмаль та глікоген, що потрапляють в організм тварин із їжею, піддаються гідролізу до мономерів (глюкози) у травній системі. У людини їхнє розщеплення починається у ротовій порожнині, продовжується у дванадцятипалій кишці і завершується утворенням глюкози біля стінок тонкого кишківника. Тут вона і всмоктується в кров, звідки може поглинатись клітинами і використовуватись у процесі гліколізу.
Ендогенні запасні полісахариди рослин (крохмаль), тварин і грибів (глікоген) включаються в гліколіз іншим шляхом. Вони зазнають не гідролізу, а фосфоролізу. Його здійснюють ферменти фосфорилаза крохмалю та глікогенфосфорилаза відповідно. Вони каталізують атаку фосфатної кислоти на глікозидний α1→4 зв’язок між останнім та передостаннім залишками глюкози з нередукуючого кінця. Продуктом реакції є глюкозо-1-фосфат. Глюкозо-1-фосфат перетворюється фосфоглюкомутазою на глюкозо-6-фосфат, який є проміжним метаболітом гліколізу. Механізм такого перетворення схожий на ізомеризацію 3-фосфогліцерату до 2-фосфогліцерату. Фосфороліз внутрішньоклітинних полісахаридів вигідний тим, що дозволяє зберегти частину енергії глікозидних зв’язків завдяки утворенню фосфорильованого моносахариду. Таким чином заощаджується одна молекула АТФ на одну молекулу глюкози.
Дисахариди[ред. | ред. код]
Як і полісахариди, дисахариди перед всмокту
Глікемія — Вікіпедія
Матеріал з Вікіпедії — вільної енциклопедії.
Глікемія (від дав.-гр. γλυκύς — «солодкий» та αἷμα — «кров») — вміст глюкози в крові. Норма для дітей до 14 років — 3,33-5,55 ммоль/л, для осіб у віці 14-60 років — 3,89-5,83 ммоль/л, у віці 60-70 років — 4,44-6,38 ммоль/л, старше 70 років — 4,61-6,10 ммоль/л[1][2]. У крові здорової людини в постабсорбтивному стані, що відбувається зранку, натщесерце, через декілька годин після прийому їжі, нормальна концентрація глюкози — 60-100 мг% або 3,3-5,5 ммоль/л[3]. Глікемія — одна з найважливіших керованих змінних в живих організмах (гомеостаз). Термін був запропонований французьким фізіологом Клодом Бернаром (1813–1878).
Фізіологічна регуляція[ред. | ред. код]
Глікемія управляється кількома фізіологічними процесами. Рівень глюкози підвищується після прийому їжі, через шлункове і кишкове поглинання легкозасвоюваних вуглеводів (низької молекулярної маси) з їжі або шляхом розщеплення з інших продуктів харчування, таких як крохмаль та полісахариди. Рівень глюкози знижується в результаті катаболізму, особливо при підвищенні температури, при фізичному навантаженні, стресі.
Інші шляхи регуляції рівня глікемії — глюконеогенез та глікогеноліз. Глюконеогенез — процес утворення в печінці і почасти в кірковій речовині нирок молекул глюкози з молекул інших органічних сполук, наприклад, вільних амінокислот, молочної кислоти, гліцерину. При глікогенолізі накопичений глікоген печінки і скелетних м’язів перетворюється на глюкозу шляхом кількох метаболічних ланцюгів.
Надлишок глюкози перетворюється на глікоген або на тригліцериди для акумулювання енергії. Глюкоза — найважливіше джерело метаболічної енергії для більшості клітин, особливо для деяких клітин (наприклад, нейрони та еритроцити), які майже повністю залежать від рівня глюкози. Мозок вимагає досить стійкої глікемії, щоб функціонувати. Концентрації глюкози в крові людини менше, ніж 3 ммоль/л, або більше, ніж 30 ммоль/л можуть призвести до втрати свідомості, судом та коми.
До регулювання метаболізму глюкози залучені кілька гормонів, таких як інсулін, глюкагон (виділяється підшлунковою залозою), адреналін (секретується залозами), глюкокортикоїди та стероїдні гормони (секретуються гонадами і залозами).
Для клінічної практики є 2 способи виявлення глікемії:
- Глікемія натще — вимірюється концентрація глюкози після 8-годинного голодування
- Тест толерантності до глюкози — триразове вимірювання концентрації глюкози крові з 30-ти хвилинним інтервалом після вуглеводного навантаження.
У деяких станах рекомендується моніторинг концентрації глюкози в крові, який зазвичай проводиться пацієнтом самостійно за допомогою портативного глюкометра.
При ряді захворювань і деяких станах концентрація глюкози в крові може або підвищуватися (цукровий діабет) — цей стан називається гіперглікемія, або знижуватися (невірно підібрана доза інсуліну при цукровому діабеті, сувора дієта, великі фізичні навантаження) — це називається гіпоглікемія.
- ↑ Рівень глюкози в крові. medictest.net. Процитовано 2016-03-11.
- ↑ Глюкоза | Анализы на EUROLAB. www.eurolab.ua. Процитовано 2016-03-11.
- ↑ Губський, Юрій (2000). БІОЛОГІЧНА ХІМІЯ. Укрмедкнига. ISBN 966-7364-41-0.
Глікозиди — Вікіпедія
Глікози́ди (від грец. γλυκύς — «солодкий» та είδο — «вигляд») — природні органічні речовини, сполуки складної будови, дуже поширені в рослинному світі; з погляду хімії — продукти конденсації циклічних форм вуглеводів (моно- або олігосахаридів) та компонента невуглеводної природи (аглікону), яким можуть бути стероїди, феноли або алкалоїди. За хімічними характеристиками подібні до гарбників.
Більшість глікозидів мають гіркий смак і специфічний запах, деякі з них володіють токсичною дією.
Молекула глікозиду складається з двох частин: цукру, він зазвичай представлений моносахаридом, і аглікону («не цукру»).[1] Маючи таким чином половину молекули цукру, вони близькі до вуглеводів, і це дає змогу об’єднати їх в одну групу.[2]
Глікозидна частина молекули, яка є циклічною формою цукрів, пов’язана через атом кисню, сірки чи азоту з радикалом органічної сполуки, що не становить собою цукру. Цей радикал називають агліконом, або геніном. Аглікони глікозидів за хімічною будовою бувають аліфатичними, ароматичними і гетероциклічними сполуками.[3] При гідролізі глікозиди розщеплюються на вуглеводневу частину (глікон) і невуглеводневу (аглікон)[4].
Інша половина молекули (аглікон) визначає специфічні особливості глікозиду, зокрема його лікувальні властивості[2]. Уважають, що кращим проникненням лікувального аглікону в клітини глікозиди завдячують цукровій частині молекули[2]. Так, аглікон у чистому вигляді, без цукру, може й не проникнути в кров і не виявити необхідної лікувальної дії. Таким чином, цукор ніби несе на собі гірку, їдку чи чимось неприйнятну частину.
Глікозиди в рослинах виникають для знешкодження всередині рослинного організму дуже активного аглікону, який міг утворитися в процесі метаболізму або потрапити ззовні.
Рослини, як відомо, в процесі фотосинтезу утворюють цукор, тому в цукрах у них ніколи дефіциту немає. Якісь надто активні, непотрібні в цей час рослині сполуки пов’язуються з цукром і стають через це для рослини нешкідливими, набувають здатності пересуватись по рослині подібно до цукрів[2]. Еволюція використала наявність глікозидів і як захист проти поїдання рослинної маси тваринами, бактеріями чи грибами, бо в них глікозиди розщеплюються, цукор засвоюється, а аглікон діє частіше як отрута.
В рослинах, що містять глікозиди, одночасно є ферменти, що їх розщеплюють на цукор і аглікон. Тому
такі рослини треба сушити якомога швидше і при температурах, що не перевищують 60 °С, після чого зберігати в сухих, добре провітрюваних приміщеннях.
Розрізняють О-глікозиди, S-глікозиди (тіоглікозиди) та N-глікозиди. Найпоширеніші у природі О-глікозиди[3].
У медицині знайшли застосування такі основні групи цих сполук:
- а) фенілглікозиди, що містять фенільний радикал в агліконі. До них належать одноатомні і багатоатомні феноли;
- б) антраглікозиди, які містять аглікон, похідний антрахінону. Їх виявлено в різних видах крушини, ревеню, алое;
- в) флавонові глікозиди, аглікон яких є похідним флавону. Переважна більшість флавонових глікозидів (рутин, катехіни) мають Р-вітамінну активність;
- г) стероїдні глікозиди, або серцеві, що належать до групи О-глікозидів, аглікони яких мають стероїдну структуру і характеризуються значною дією на серцевий м’яз. Глікозиди конвалії травневої (звичайної), горицвіту весняного, наперстянки мають давню традицію застосування в народній, а згодом і науковій медицині;
- д) S-глікозиди, або тіоглікозиди, — найменш поширена група глікозидів серед рослинного світу. Їх знайдено в насінні сарепської і чорної гірчиці, редьці посівній, хроні звичайному, капусті. Названі сполуки гострі, пекучі на смак. Вони збуджують апетит, виявляють місцеву подразнюючу дію, завдяки чому посилюють кровообіг при зовнішньому застосуванні, справляють активний бактерицидний і бавктеріостатичний влив на патогенні групи мікроорганізмів, що викликають запалення шкіри, підшкірної основи та м’язів людини.[3]
Специфічна дія глікозиду зумовлена типом аглікону. Це здебільшого кристалічні сполуки, майже завжди гіркі на смак, мають специфічний запах. Беруть участь у процесах обміну речовин.
Деякі важливі приклади[ред. | ред. код]
Особливо багаті на глікозиди рослини, але виявлені вони і в організмах тварин. У рослинах найчастіше зустрічаються такі глікозиди:
Для рослин з родини капустяних (хрестоцвітих) характерна наявність у глікозидах сірковмісних речовин. Це — глікозиди синігрин і синальбін у насінні гірчиці[2].
У розоцвітих поширений амигдалін, який розпадається з виділенням синильної (ціанової) кислоти. Він є в насінні гіркого мигдалю, абрикоса, вишень, персика, сливи, лавровишні, в квітках і листі черемхи, а також зустрічається в представників зовсім інших родин[2].
У зелених частинах льону глікозид лінамарин також містить синильну кислоту — дуже сильну отруту[2].
У жовтецевих зустрічається анемонін і протоанемонін.
Строфантин характерний для рослин родини кутрових — строфанту (чилібухи), олеандра, кендирю коноплевидного.
До групи глікозидів в організмі тварин і людини належать цереброзиди мозку і нуклеозиди. Деякі глікозиди (стрептоміцин, азонін, строфантин) застосовують у медицині.
Глікозиди, як і багато інших органічних речовин, можна одержувати штучним шляхом, однак при цьому утворюються численні ізомери, тотожні молекули, з дещо іншим розміщенням частин, які, проте, не мають фізіологічної чи фармакологічної активності. Розділити таку суміш з практично однакових у хімічному відношенні молекул на активні й неактивні дуже важко, часом просто неможливо, бо число ізомерів іноді налічує 128, 256 і більше варіантів, а тому для виготовлення ліків потрібна природна рослинна сировина. Її заготовляють у природі або ж вирощують на полях.
- ↑ Скурихин И. М., Нечаев А. П. Все о пище с точки зрения химика: Справ. издание. — М.: Высш. шк. 1991. — 288 с. ISBN 5-06-000673-5 (рос.)
- ↑ а б в г д е ж Лікарські рослини: Енциклопедичний довідник/ Відп. ред. А. М. Гродзінський. — К.: Видавництво «Українська енциклопедія» ім. М. П. Бажана, Український виробничо-комерційний центр «Олімп», 1992. — 544с. ISBN 5-88500-055-7
- ↑ а б в г Товстуха Є. С. Фітотерапія. — К.: Здоров’я, 1990.-304 с., іл., 6,55 арк. іл. ISBN 5-311-00418-5
- ↑ а б Канюка О. І., Скорохід В. Й., Гуфрій Д. Ф. Клінічна ветеринарна фармакологія. — К: В-во УСГА, 1993. — 293 с. ISBN 5-7987-0067-4
- Глосарій термінів з хімії // Й. Опейда, О. Швайка. Ін-т фізико-органічної хімії та вуглехімії ім. Л. М. Литвиненка НАН України, Донецький національний університет. — Донецьк : Вебер, 2008. — 758 с. — ISBN 978-966-335-206-0
Глюкоза — Википедия
Материал из Википедии — свободной энциклопедии
| Глюкоза | |
 | |
 | |
| Общие | |
|---|---|
| Систематическое наименование | (2R,3S,4R,5R)-2,3,4,5,6-пентагидроксигексаналь (D-глюкоза), (2S,3R,4S,5S)-2,3,4,5,6-пентагидроксигексаналь (L-глюкоза) |
| Традиционные названия | Глюкоза, глюкогексоза |
| Хим. формула | С6H12O6 |
| Физические свойства | |
| Молярная масса | 180,16 г/моль |
| Плотность | 1.54-1,60 г/см³ |
| Термические свойства | |
| Т. плав. | α—D-глюкоза: 146 °C β—D-глюкоза: 150 °C |
| Классификация | |
| Рег. номер CAS | 50-99-7 (D-глюкоза) 921-60-8 (L-глюкоза) |
| Рег. номер EINECS | 200-075-1 |
| RTECS | LZ6600000 |
| ChEBI | 17234 |
| Приводятся данные для стандартных условий (25 °C, 100 кПа), если не указано иного. | |
Глюко́за, или виноградный сахар, или декстроза (D-глюкоза), С6H12O6 —
органическое соединение, моносахарид (шестиатомный гидроксиальдегид, гексоза), один из самых распространённых источников энергии в живых организмах на планете[1]. Встречается в соке многих фруктов и ягод, в том числе и винограда, от чего и произошло название этого вида сахара. Глюкозное звено входит в состав полисахаридов (целлюлоза, крахмал, гликоген) и ряда дисахаридов (мальтозы, лактозы и сахарозы), которые, например, в пищеварительном тракте быстро расщепляются на глюкозу и фруктозу.
Физические свойства
Бесцветное кристаллическое вещество без запаха. Обладает сладким вкусом, растворимо в воде, в реактиве Швейцера (аммиачном растворе гидроксида меди [Cu(NH3)4](OH)2), в концентрированном растворе хлорида цинка и концентрированном растворе серной кислоты.
В 2 раза менее сладкая, чем сахароза.
Строение молекулы
Глюкоза может существовать в виде циклов (α- и β-глюкозы) и в виде линейной формы (D-глюкоза).
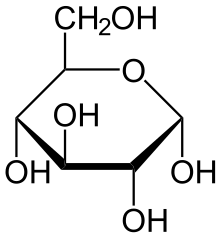
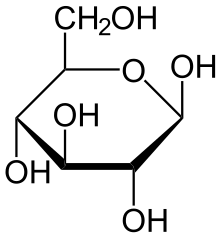
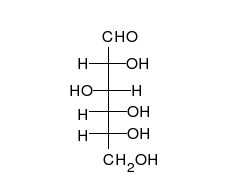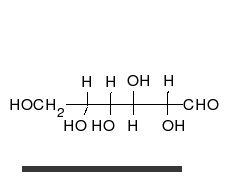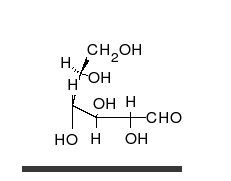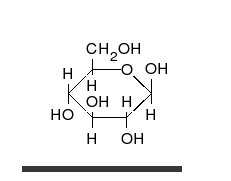
Глюкоза — конечный продукт гидролиза большинства дисахаридов и полисахаридов.
Получение
В промышленности глюкозу получают гидролизом крахмала и целлюлозы.
В природе глюкоза образуется растениями в процессе фотосинтеза.
Химические свойства
Глюкоза может восстанавливаться в шестиатомный спирт (сорбит). Глюкоза легко окисляется. Она восстанавливает серебро из аммиачного раствора оксида серебра и медь(II) до меди(I).
Проявляет восстановительные свойства. В частности, в реакции растворов сульфата меди(II) с глюкозой и гидроксидом натрия. При нагревании эта смесь реагирует с обесцвечением (сульфат меди сине-голубой) и образованием красного осадка оксида меди(I).
Образует оксимы с гидроксиламином, озазоны с производными гидразина.
Легко алкилируется и ацилируется.
При окислении образует глюконовую кислоту, если воздействовать сильными окислителями на её гликозиды, и гидролизовав полученный продукт можно получить глюкуроновую кислоту, при дальнейшем окислении образуется глюкаровая кислота.
Биологическая роль
Глюкоза — основной продукт фотосинтеза, образуется в цикле Кальвина.
В организме человека и животных глюкоза является основным и наиболее универсальным источником энергии для обеспечения метаболических процессов. Глюкоза является субстратом гликолиза, в ходе которого она может окислиться либо до пирувата в аэробных условиях, либо до лактата в случае анаэробных условий. Пируват, полученный таким образом в гликолизе, далее декарбоксилируется, превращаясь в ацетил-КоА (ацетилкоэнзим А). Также в ходе окислительного декарбоксилирования пирувата восстанавливается кофермент НАД+. Ацетил-КоА далее используется в цикле Кребса, а восстановленный кофермент используется в дыхательной цепи.
Глюкоза депонируется у животных в виде гликогена, у растений — в виде крахмала, полимер глюкозы — целлюлоза является основной составляющей клеточных оболочек всех высших растений.
У животных глюкоза помогает пережить заморозки. Так, у некоторых видов лягушек перед зимой повышается уровень глюкозы в крови, за счет чего их тела способны выдержать заморозку во льду.
Применение
Глюкозу используют при интоксикации (например, при пищевом отравлении или деятельности инфекции), вводят внутривенно струйно и капельно, так как она является универсальным антитоксическим средством. Также препараты на основе глюкозы и сама глюкоза используются эндокринологами при определении наличия и типа сахарного диабета у человека (в виде стресс-теста на ввод повышенного количества глюкозы в организм).
Ссылки
Примечания
| Общие: | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Геометрия | |||||||||||||||
| Моносахариды |
| ||||||||||||||
| Мультисахариды | |||||||||||||||
| Производные углеводов | |||||||||||||||
Целюлоза — Вікіпедія
Матеріал з Вікіпедії — вільної енциклопедії.
Целюло́за, (С6Н10О5)x — природний полімер, полісахарид, волокниста речовина, головна складова частина оболонки рослинних клітин. У найбільшій кількості міститься у деревині, волокнах бавовни, льону та ін.
Целюлоза являє собою лінійний стереорегулярний (синдіотактичний) природний полісахарид, побудований з ангідридів D-глюкопіранози. Стереорегулярна будова макромолекули й стійкість конфірмаційної форми її елементарної ланки виділяє целюлозу із усього ряду полісахаридів, у тому числі й найбільшою стійкістю до хімічних впливів.
Целюлоза була відкрита 1838 року французьким хіміком Ансельмом Паєном.
Величина Х у молекулах целюлози завжди становить приблизно 3 000, але може досягати від 6 000 до 12 000. Найчистіша природна целюлоза — бавовняне волокно — становить 85—90% целюлози. У деревині хвойних дерев приблизно 50% целюлози (до складу деревини нарівні з целюлозою входять її супутників, серед них найважливішим є лігнін — природний полімер, побудований із декількох ароматичних сполук ряду бензолу, і геміцелюлози — споріднені з целюлозою полісахариди).
Целюлоза не розчиняється у воді і діетиловому ефірі і етиловому спирті. Вона не розщеплюється під дією розбавлених кислот, стійка до дії лугів і слабких окисників. При обробці на холоді концентрованою сірчаною кислотою целюлоза розчиняється в ній, утворюючи в’язкий розчин. Якщо цей розчин вилити в надлишок води, виділяється білий пластевидний продукт, так званий амілоїд, що являє собою частково гідролізовану целюлозу. Він схожий з крохмалем за реакцією з йодом (синє забарвлення; целюлоза не дає цієї реакції). Якщо непроклеєний папір опустити на короткий час в концентровану сірчану кислоту і потім терміново промити, то амілоїд, що утвориться, склеїть волокна паперу, роблячи його щільнішим та міцнішим. Так виготовляється пергаментний папір.
Під дією концентрованих розчинів мінеральних кислот і підігріву целюлоза піддається гідролізу, кінцевим продуктом якого є глюкоза.
Значення целюлози дуже велике, достатньо вказати, що велика кількість бавовняного волокна йде на виробництво бавовняних тканин. З целюлози виробляють папір і картон, а способом хімічної переробки — цілий ряд різноманітних продуктів: штучне волокно, пластичні маси, лаки, бездимний порох, етиловий спирт.
У 2000 р. було вироблено 171 млн т целюлозної пульпи і 323 млн т паперових виробів.[3]
Використовується як наповнювач деяких пластмас, у вигляді волокон, упаковочного матеріалу, фармацевтичній практиці.[4] Важливими штучними полімерами є прості і складні ефіри целюлози (продукти заміщення атомів водню в ОН-групах целюлози вуглеводневими (-R) або кислотними (-COR) радикалами): ацетат целюлози (ацетилцелюлоза), ацетопропіонат целюлози, гідроксиетилцелюлоза, карбоксиметилцелюлоза (КМЦ), Na-карбоксиметилцеоюлоза, метилцелюлоза, метилетилцелюлоза, нітроцелюлоза, целулоїд (целон), етроли.[4] Оксицелюлозу ввели в застосування до 1950-х років для обміну іонів з білками і пептидами. Пізніше були синтезовані складніші форми: карбоксиметилцелюлоза, диетиаміноетанцелюлоза, сульфометилцелюлоза, сульфоетилцелюлоза, триетиламіноетанцелюлоза.[5]
Їстівні властивості[ред. | ред. код]
Целюлоза як і крохмаль є одним з видів збереження глюкози. Людський шлунок не здатен розкладати її волокна. Однак целюлоза, що міститься у рослинній їжі, надає їй об’єм, що стимулює перистальтику кишківника.[6] Її іноді додають до складу кормів для тварин.[7]
- ↑ Nishiyama, Yoshiharu; Langan, Paul; Chanzy, Henri (2002). Crystal Structure and Hydrogen-Bonding System in Cellulose Iβ from Synchrotron X-ray and Neutron Fiber Diffraction. J. Am. Chem. Soc 124 (31): 9074–82. PMID 12149011. doi:10.1021/ja0257319. .
- ↑ Cellulose gel
- ↑ World Agriculture — Towards 2015—2030. An FAO prespective… FAO. Forestry.pdf.
- ↑ а б Шефтель В. О. Вредные вещества в пластмассах. — М.: Химия, 1991—574 с. ISBN 5-7245-0590-8
- ↑ Растительный белок/Пер. с фр. В. Г. Долгополова; Под ред. Т. П. Микулович. — М.: Агропромиздат, 1991. — 684 с.: ил. ISBN 5-10-001276-5 (СССР) ISBN 2-85206-239-9 (Франция) (с.:74)
- ↑ Введение в биологию. П.Кемп, К.Армс // Мир, М.1988 — 671 с. — С.244
- ↑ Яка особливість целюлози в складі кормів.
Глюкоза — Уикипедия
Уикипедия — ашық энциклопедиясынан алынған мәлімет
Глюкоза (жүзім қанты) — Денеге тез және жеңіл сіңетін қанттың қарапайым түрі. Ол жемісжидектерде және балда болады, денедегі суды қалпына келтіруге пайдалануға арналған ақ ұнтақ түрінде сатылады. Глюкоза — моносахарид тобындағы көмірсутек. Тамақ өнеркәсібінде, медицинада және аналитикалық химияда реактив ретінде қолданылады. Өсімдік жемістерінде және жануар организмдерінде болатын жүзім қанты. Сахароза, лактоза құрамына кіреді және крахмал, гликоген, целлюлоза түзеді. Глюкоза маңызды зат алмасу азығы болып есептеледі, тірі тіндерді энергиямен қамтамасыз етеді. Адам организмде қандағы глюкозаның керекті мөлшері (100 мг % шамасында) гликогеннің синтезі мен ыдырауы арқылы тұрақтандырылады. Өндірісте Глюкоза крахмалды гидролиздеу арқылы алынады.[1]
Глюкозаның табиғатта түзілуі мен алынуы[өңдеу]
- 1. Табиғатта глюкоза фотосинтез реакциясы нәтижесінде түзіледі.
- 2. Қантты заттарды, ең алғаш A.M. Бутлеров (1861 ж.) кальций гидроксиді қатысында формальдегидтен синтездеді:
- 3. Өнеркәсіпте глюкозаны, негізінен, полисахаридтерді (крахмалды немесе целлюлозаны) гидролиздеп алады:
Физикалық қасиеттері[өңдеу]
Түссіз, қатты кристалды зат, дәмі тәтті, суда жақсы ериді.
Химиялық қасиеттері[өңдеу]
Глюкоза құрамында альдегид тобы бар көпатомды спирт болғандықтан, альдегидтерге және көпатомды спирттерге тән екі жақты қасиет көрсетеді.
Альдегид тобына тән реакциялары[өңдеу]
- 1. Глюкоза жақсы тотықсыздандырғыш, жұмсақ тотықтырғыштардың әсерінен, мысалы, күміс оксидінің аммиактағы ерітіндісімен альдегид тобы ғана тотығып, глюкон қышқылына айналады:
- 2. Мыс (II) гидроксидімен қыздырғанда, альдегид тобы тотығып, карбоксил тобына айналады және мыс (І) оксидінің қызыл түсті тұнбасы түзіледі:
- 3. Глюкозаның альдегид тобы тотықсыздану реакциясына да түседі. Катализатор қатысында сутекпен тотықсызданғанда, глюкоза алтыатомды спирт — сорбит түзеді:
Глюкозаның көпатомды спирттерге тән қасиеттері[өңдеу]
- 4. Бөлме температурасында мыс (II) гидроксидімен әрекеттескенде, ашық көк түсті ерітінді түзеді, бұл глюкозаның көпатомды спирт екенін дәлелдейді. Глюкоза көпатомды спирт болғандықтан, күрделі және жай эфирлер түзеді.
- 5.Күрделі эфирлер түзілуі. Қанттардың гидроксил топтары этерификация реакциясына оңай түседі. Кең таралған этерификация реакциясы ацилдеу, оны сірке ангидридімен катализатор (ZnCl2, H2SO4 натрий ацетаты) қатысында жүргізеді. Гексозалардың тұйық құрылымы тұйық пентаацетаттар түзеді. Қышқылмен әрекеттесу реакциясы глюкозаның құрамында бес гидроксил тобы бар екенін дәлелдейді. Мысалы, глюкоза сірке ангидридімен мырыш хлориді қатысында әрекеттесіп, α-пентаацетаттар түзеді:
6. Жай эфирлер түзілуі. Глюкозаны метил спиртімен қышқыл ортада алкилдесе, реакцияға гликозид гидроксилі түседі, реакция нәтижесінде жай эфир түзіледі. Бұл реакция глюкозаның циклді түріндегі глюкозид гидроксил тобы басқа гидроксилдермен салыстырғанда реакцияға түскіш екенін көрсетеді:
Глюкозаның маңызды химиялық қасиеттерінің бірі — әр түрлі микроорганизмдер әсерінен ашуы.
- 7. Спирттік ашуы. Ашытқы ферменті әсерінен ашып, глюкозадан этанол түзіледі. Бұл процестің химизмі өте күрделі. Жүретін өзгерістерді қысқаша былай жазады:
- С6Н12O6 → 2С2Н5ОН + 2СO2
- 8. Сүт қышқылды ашуы. Глюкоза сүт қышқылды ашығанда, сүт қышқылы түзіледі. Процесс сүт қышқылды бактерия ферменттерінің әсерінен жүреді:
Сүт қышқылы екі функционалдық тобы бар болғандықтан (гидроксил және карбоксил топтары), екі түрлі қасиет көрсетеді. Сүт қышқылы сүт іріген кезде пайда болады, оны лакмус қағазы арқылы анықтауға болады.
- 9. Тотығуы. Органикалық қосылыс болғандықтан, глюкоза толық тотыға алады. Ең соңғы өнім ретінде көміртек (IV) оксиді мен су түзіледі. Бұл процесті ықшамдап былай жазуға болады:
- С6Н12O6 + 6O2 → 6СO2 + 6Н2O + Q
Глюкозаның маңызы, қолданылуы[өңдеу]
Глюкоза адам ағзасында зат алмасу процесіне қатысады. Тамақ арқылы қабылданған крахмал, қант сияқты күрделі көмірсулар ферменттердің әсерінен ас қорыту жолдарында глюкозаға ыдырайды. Адам ағзасында глюкозаның негізгі бөлігі энергия қоры ретінде жұмсалады (28-кесте). Тірі ағза клеткаларында глюкоза оттекпен бірнеше саты арқылы біртіндеп тотығады. Нәтижесінде, фотосинтез процесінде сіңірілген энергия бөлінеді:
- С6Н12O6+ 6O2 → 6СO2 + 6Н2O + Q
Глюкоза ағзада оңай қорытылып энергия бөлетіндіктен денсаулықты нығайтатын дәрі ретінде медицинада қолданады. Кондитер өнеркәсібінде (мармелад, печенье, карамель, күлше нандар жасағанда), мата өңдегенде пайдаланылады. Тотықсыздандырғыш қасиетіне байланысты айна және шырша ойыншықтарын жасағанда қолданады.
Глюкоза — бағалы қоректік зат. Глюкоза жүзімнің және басқа өсімдіктердің шырынында болады. Оның қалдықтарынан өте маңызды полисахаридтер (целлюлоза мен крахмал) түзіледі. Глюкозаның сүт қышқылдық ашу процесінің ауыл шаруашылығында маңызы зор. Мысалы, сүт қышқылды ашу арқылы айран, сүзбе, қаймақ, т.б. сүт өнімдерін алады. Капуста тұздағанда немесе шөпті сүрлемге салғанда, реакция нәтижесінде түзілген сүт қышқылы қышқылдык қасиет көрсетіп, өнімді шіруден қорғайды.
Глюкозаның спирттік ашуы да практикада, мысалы, сыра жасағанда қолданылады.[2]
Глюкоза өндіру[өңдеу]
Глюкоза өндіру — крахмалды өндірісте күкірт қышқылының ерітіндісінде қайнатып алу. Мұндай процесті қышқылды гидролиз дейді. Қайнағаннан соң қышқылды бормен бейтараптандырады, ал глюкоза ерітіндісі ерімейтін кальций сульфатынан сүзіліп, вакуумда шулықтырады. Пайда болған қою шырынды сірне деп атайды. Кристалды глюкоза алу үшін крахмалды толық гидролиздейді, қоюланған қою шырынды формаларға салып қатырады. Сусыз глюкоза 146° С-та балқиды және суда жақсы ериді. Тәттілігі сахарозадан екі еседей кем болады. Глюкозада күшті қышқылдандырғыштардың әсерінен (мысалы, Н1Ч03) екі негізді оксиқышқыл (қант қышқылы) пайда болады. Қайта қалыптасқанда қант қышқылы алты атомды спирт-сорбитке айналады.[3]
Дереккөздер[өңдеу]
- ↑ Вирусология, иммунология, генетика, молекулалық биология. Орысша-қазақша сөздік. – Алматы, «Ана тілі» баспасы, 1993 жыл. ISBN 5-630-0283-X
- ↑ Химия: Жалпы білім беретін мектептің жаратылыстану-математика бағытындағы 11-сыныбына арналған оқулық / Ә. Темірболатова, Н. Нұрахметов, Р. Жұмаділова, С. Әлімжанова. – Алматы: «Мектеп» баспасы, 2007. – 352 бет, суретті. ISBN 9965-36-092-8
- ↑ Қазақ тілі терминдер сөздігі I том
Гликолиз и глюконеогенез — знания для студентов-медиков и врачей
Гликолиз и глюконеогенез — это метаболические процессы, ответственные за деградацию глюкозы или синтез глюкозы соответственно. При гликолизе распад молекул глюкозы приводит к образованию двух чистых молекул аденозинтрифосфата (АТФ), которые обеспечивают легкодоступный источник энергии для различных реакций в клетке, и двух молекул пирувата, которые в дальнейшем могут быть преобразованы в лактат (используемый в глюконеогенезе). , ацетил-КоА (используется в цикле лимонной кислоты), оксалоацетат (используется в цикле лимонной кислоты) и аланин (используется в реакциях трансаминирования).Гликолиз, который происходит исключительно в цитоплазме, является единственным источником АТФ в клетках, в которых отсутствуют митохондрии (например, красные кровяные тельца). В глюконеогенезе глюкоза, которая обеспечивает эугликемию во время голодания, синтезируется из неуглеводных предшественников, таких как глюкогенные аминокислоты (в основном аланин и глутамин), жирные кислоты с нечетной цепью, глицерин, пируват и лактат. В то время как гликолитический путь происходит во всех клетках, глюконеогенез происходит почти исключительно в печени. Фосфофруктокиназа-1 является ферментом, ограничивающим скорость гликолиза, в то время как фруктозо-1,6-бисфосфатаза является ферментом, ограничивающим скорость глюконеогенеза.Метаболизм глюкозы в основном контролируется гормонами, такими как инсулин, который стимулирует гликолиз, и глюкагон, который стимулирует глюконеогенез. Глюкоза также может быть переведена на пентозофосфатный путь (также известный как гексозо-монофосфатный шунт), который представляет собой метаболический путь, который генерирует никотинамидадениндинуклеотидфосфат (НАДФН) и рибозо-5-фосфат из глюкозо-6-фосфата. Пентозофосфатный путь происходит исключительно в цитозоле и очень активен в коре надпочечников, печени и эритроцитах (эритроцитах).Рибозо-5-фосфат необходим для синтеза нуклеотидов, в то время как НАДФН необходим для синтеза холестерина, стероидов, восстановления глутатиона и респираторного взрыва. Глюкоза 6-
.
Glukoza — Википедия, вольна энциклопедия
 | Ten artykuł dotyczy D -glukozy. Zobacz też: L -glukoza. |
 α- D -glukopiranoza (projekcja stereochemiczna) | |||||||||||||||
| |||||||||||||||
| Ogólne informacje | |||||||||||||||
| Wzór sumaryczny | C 6 H 12 O 6 | ||||||||||||||
| Masa molowa | 180,16 г / моль | ||||||||||||||
| Wygląd | bezbarwne, drobne kryształki o słodkim smaku [1] | ||||||||||||||
| Identyfikacja | |||||||||||||||
| номеров CAS | 50-99-7 26655-34-5 (изомер α- D ) 28905-12-6 (изомер β- D ) 58367-01-4 (рацемат) 77938-63-7 ( моногидрат) | ||||||||||||||
| PubChem | 5793 | ||||||||||||||
| Лекарственный банк | DB02379 | ||||||||||||||
| |||||||||||||||
| Podobne zwizki | |||||||||||||||
| Podobne związki | фруктоза, сорбоза, галактоза и инне хексозы | ||||||||||||||
| Pochodne | глюкозо-6-фосфоран | ||||||||||||||
| Jeżeli nie podano inaczej, dane dotyczą stanu standardowego (25 ° C, 1000 гПа) | |||||||||||||||
| Klasyfikacja medyczna | |||||||||||||||
| УВД | V04CA02, B05CX01, V06DC01, C05BB56 | ||||||||||||||
| |||||||||||||||
| |||||||||||||||
Glukoza , D -glukoza (stgr.γλυκός glykós «słodki», -oza przyrostek oznaczajcy węglowodany) — органический химический, моносахарид (cukier prosty) z grupy aldoheksoz. Jest białym, drobnokrystalicznym ciałem stałym [1] , z roztworów wodnych łatwo krystalizuje jako monohydrat [7] . Jest bardzo dobrze rozpuszczalna w wodzie [1] (nie zmienia pH roztworu). Ma słodki smak, nieco mniej интенсивы од сахарозы [8] .
Enancjomerem D -glukozy jest L -glukoza, niewystępująca w organmach wyższych.
Glukoza powstaje w organizationmach roślinnych podczas fotosyntezy [9] :
- 6H 2 O + 6CO 2 + hν (energia świetlna) → C 6 H 12 O 6 + 6O 2 ↑; ΔE = −2872 кДж / моль (–687 ккал / моль)
- gdzie: h — stała Plancka; ν — częstotliwość fali
Występuje w znacznych ilościach w owocach (szczególnie w winogronach, stąd nazywana jest czasem cukrem iiodzime ).W organizationmach jest składowana w postaci dwóch polimerów: skrobi u roślin i glikogenu u zwierząt.
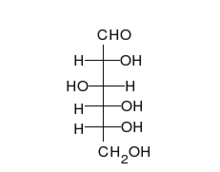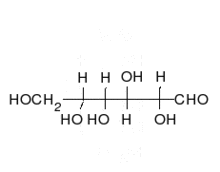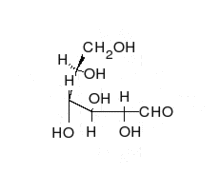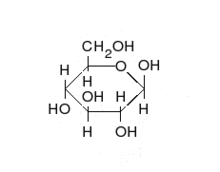
Krystaliczna glukoza ma budowę cykliczną. Sześcioczłonowy pierścień hemiacetalu powstaje wyniku reakcji grupy aldehydowej przy atomie węgla C1 z grupą hydroksylową przy atomie węgla C5 w łańcuchu. Możliwe są zatem dwa anomery — α oraz β w zależności od położenia grupy OH przy pierwszym atomie węgla formy cyklicznej.
W roztworze wodnym oba anomery mogą ulegać reakcji otwarcia pierścienia i ponownego zamknięcia z utworzeniem ponownie takiego samego lub przeciwnego anomeru.Proces taki nazywa się mutarotacją. Forma otwartołańcuchowa występuje wyłącznie w roztworze wodnym, a jej termodynamiczna stableność jest znacznie mniejsza od formy cyklicznej, dlatego równęysza od formy cyklicznej, dlatego równęysza od formy cyklicznej, dlatego równęysza od formy cyklicznej, dlatego równęycy cyęstęjjjkzówy przesuni.
W łańcuchowej formie glukozy znajdują się cztery asymetryczne atomy węgla (tj. C2, C3, C4 и C5), podczas gdy w formie cyklicznej występuje pięč takich atomcówny asyodatry.W zależności od położenia grupy karbonylowej w łańcuchu i konfiguracji asymetrycznych atomów węgla, możliwych jest kilkadziesiąt izomerów glukozy (зоб. Heksozy). Jednym z często występujących naturalnie izomerów glukozy jest fruktoza, różniąca się połoeniem grupy karbonylowej.
Właściwości chemiczne [edytuj | эдитуй код]
Naturalna glukoza jest jednym z wielu izomerów optycznych tego związku — α- D -glukopiranozą (dekstrozą). Jest ona czynna optycznie — w temperaturze pokojowej skręca płaszczyznę spolaryzowanego światła białego w prawo.Skręcalność właściwa anomeru α wynosi + 112 °, natomiast anomeru β + 19 °. W roztworze wodnym w wyniku mutarotacji ustala się stan równowagi obu anomerów w stosunku 36:64, o skręcalności właściwej + 52,6 ° [10] .
Glukoza w przeciwieństwie do fruktozy odbarwia wodę bromową w obecności kwaśnego węglanu sodu [11] (następuje utlenienie grupy aldehydowej do grupy karboksylowej, grupy karboksylowej, 1, 9015).
- CH 2 OH− (CHOH) 4 −CHO + Br 2 + 2NaHCO 3 → CH 2 OH− (CHOH) 4 −COOH + 2NaBr + H 2 O + 2CO 2 ↑
Pod wpływem kwasu azotowego grupa aldehydowa oraz atom węgla C6 są utleniane do grup karboksylowych, w wyniku czego powstaje kwas glukarchodowy 90 [kwas grupa grupa].
W temperaturze powyżej 150 ° C topi się i brunatnieje, tworząc (podobnie jak sacharoza) karmel [1] .
Właściwości biologiczne [edytuj | эдитуй код]
D -Glukoza jest podstawowym związkiem energetycznym dla większości organizationmów, przechowywanym pod postaci polimerów — skrobi i glikogenu. Stanowi cukier najłatwiej przyswajalny przez człowieka [1] . W processie glikolizy jest rozkładana na kwas pirogronowy. Jest także substratem wielu processów zachodzących w komórce, a u roślin, jako celuloza, podstawowym budulcem ściany komórkowej.
Открытие глюкозы в розлив [едитуй | эдитуй код]
 | Ta sekcja od 2011-10 wymaga zweryfikowania podanych informacji. Należy podać wiarygodne źródła, najlepiej w formie przypisów bibliograficznych. Część lub nawet wszystkie informacje w sekcji mogą być nieprawdziwe. Яко pozbawione źródeł mogą zostać zakwestionowane i usunięte. Dokładniejsze informacje o tym, co należy poprawić, być może znajdują się w dyskusji tej sekcji. Po wyeliminowaniu niedoskonałości należy usunąć szablon {{Dopracować}} z tej sekcji. |
Glukozę, podobnie jak inne cukry redukujące, można wykryć m.in. za pomocą prób Trommera i Tollensa [1] . W obydwu próbach glukoza jest przeprowadzana w kwas glukonowy.
Próba Trommera [edytuj | эдитуй код]
Do świeżo strąconego osadu wodorotlenku miedzi (II) dodaje się badany roztwór i ogrzewa. W obecności glukozy niebieski osad Cu (OH) 2 zmienia barwę na ceglastoczerwoną, wskutek redukcji do Cu 2 O:
- 2Cu (OH) 2 + C 6 H 12 O 6 → Cu 2 O + C 6 H 12 O 7 + 2H 2 O
Próba Tollensa [edytuj | эдитуй код]
Do probówki napełnionej niewielką ilością roztworu azotanu srebra należy dodać kilka kropli stężonego roztworu wodorotlenku sodu (NaOH), a następnie dolewać kroplami powelk ilością roztworu azotanu srebra należy dodać kilka kropli stężonego roztworu wodorotlenku sodu (NaOH), a następnie dolewać kroplami rozénés de la roztworu, wodorotlenku sodu (NaOH), a następnie dolewać kroplami rozizakuzeraworu.
1. AgNO 3 + NaOH → AgOH + NaNO 3
2.a) 2 AgOH → Ag 2 O + H 2 O
2.b) Ag 2 O + CH 2 (OH) (CHOH) 4 CHO → 2 Ag + CH 2 (OH) (CHOH) 4 COOH
Tak przygotowaną probówkę należy umieścić w zlewce z gorącą wodą (na łaźni wodnej). Na koniec do probówki dodaje się badany roztwór.
Podczas ogrzewania subcji o barwie brunatnej — Ag 2 O (powstałej z reakcji AgNO 3 z NaOH) — na ściankach probówki osadza się warstewka zerameticznego srebónsrebónszrebón zeb .
3. Ag 2 O + глюкоза → 2 Ag + квас-глюконозный (квас-глюконовый)
Ag 2 O + C 6 H 12 O 6 → 2 Ag + C 6 H 12 O 7
W medycynie (np. Kroplówki), a także w przemyśle spożywczym (np. Syrop glukozowy [12] , syrop glukozowo-fruktozowy) oraz farmaceutycznym [1] .
- ↑ a b c d e f g h i j k Podręczual s.Hassa (красный), JanuszJ. Mrzigod (красный), JanuszJ. Новаковский (ред.), Катовице: Видеограф II, 2004, с. 150–151, ISBN 83-7183-240-0.
- ↑ a b Farmakopea Polska VIII , Polskie Towarzystwo Farmaceutyczne, Warszawa: Urząd Rejestracji Produktów Leczniczych, Wyrobów Medycznych i Produktzych i Produktzych Biobójc. 3491, ISBN 978-83-88157-53-0.
- ↑ a b c d Glukoza ( угл. ). [martwy link] Химическая база данных.Wydział Chemii Uniwersytetu w Akronie. [dostęp 03.09.2012]. [niewiarygodne ródło?]
- ↑ a b c d e Glukoza ( угл. ). [martwy link] Химическая база данных. Wydział Chemii Uniwersytetu w Akronie. [dostęp 03.09.2012]. [niewiarygodne ródło?]
- ↑ Глюкоза , [w:] ChemIDplus [онлайн], Национальная медицинская библиотека США [dostęp 2012-09-03] ( англ.).
- ↑ Glukoza (nr G8270) — карта характеристик продукта Sigma-Aldrich (Merck KGaA) на obszar Polski.
- ↑ Р. К. Дж. Киллиан, В. Г. Ферриер, Д. В. Янг. Предварительное исследование кристаллической и молекулярной структуры α- D -глюкозы моногидрата . «Acta Crystallographica». 15 (9), с. 911–912, 1962. DOI: 10.1107 / S0365110X62002352 ( угл. ).
- ↑ Обучение, Ресурсы Пищи. Государственный университет Орегона, Корваллис, штат Орегон. [dostęp 17.05.2010].[zarchiwizowane z tego adresu (2015-01-09)].
- ↑ Теодор Браун, Джон Д. Нельсон, Кеннет В. Кемп: Химия. Центральная Наука . Верхняя Сэдл-Ривер, штат Нью-Джерси: Prentice Hall, 2003, с. 958. ISBN 0-13-048450-4.
- ↑ John McMurry: Chemia organiczna . Wyd. 3. Т. 4. Варшава: PWN, 2005, с. 954–955. ISBN 83-01-14406-8.
- ↑ Utlenienie cukrów. W: Александр Kołodziejczyk: Naturalne związki organiczne . Wyd. 2 попр. Варшава: Wydawnictwo Naukowe PWN, 2006, с.250–253. ISBN 978-83-01-14316-9.
- ↑ Czym jest syrop glukozowy i z czego jest wytwarzany? , [w:] Food-Info.net [онлайн], Университет Вагенингена, Голландия [dostęp 09.01.2016].
| V06D — Inne preparaty żywieniowe | |||||||||||||||||||||||||||||||||||
.
Мальабсорбция глюкозы и галактозы — Genetics Home Reference
Мутации в гене SLC5A1 вызывают мальабсорбцию глюкозы и галактозы. Ген SLC5A1 предоставляет инструкции по производству белка, называемого протеином котранспортера натрия / глюкозы 1 (SGLT1). Этот белок содержится в основном в кишечнике и почках. Он охватывает мембрану клеток в этих системах организма и перемещает (транспортирует) глюкозу и галактозу из-за пределов клетки внутрь клетки. Натрий и вода транспортируются через клеточную мембрану вместе с сахарами в этом процессе.Глюкоза и галактоза — простые сахара; они присутствуют во многих продуктах питания или могут быть получены при расщеплении лактозы или других сахаров и углеводов в рационе во время пищеварения.
В кишечном тракте белок SGLT1 помогает организму усваивать глюкозу и галактозу из пищи, чтобы организм мог их использовать. Во время переваривания пищи белок переносит сахар в клетки, выстилающие стенку кишечника (клетки кишечного эпителия), по мере прохождения пищи.
Белок SGLT1 в клетках почек играет роль в поддержании нормального уровня глюкозы в крови.Почки фильтруют отходы из крови и выводят их с мочой. Они также реабсорбируют необходимые питательные вещества и возвращают их в кровь. Белок SGLT1 транспортирует глюкозу в специализированные клетки почек, гарантируя, что сахар возвращается в кровоток и не попадает в мочу.
Мутации гена SLC5A1 нарушают или исключают функцию белка SGLT1. В результате глюкоза и галактоза не абсорбируются эпителиальными клетками кишечника, а вместо этого накапливаются в кишечном тракте.Кроме того, вода, которая обычно транспортировалась бы вместе с сахарами, остается в кишечном тракте, что приводит к обезвоживанию тканей организма и тяжелой диарее. Белок SGLT1 в клетках почек не может транспортировать глюкозу; однако другие белки в почках способны абсорбировать достаточное количество глюкозы в кровоток, так что глюкозурия у людей с мальабсорбцией глюкозы-галактозы бывает легкой, если вообще присутствует.
.
Контроль диабета и глюкозы — Продление жизни
Связаться с нами
0
дополнительных товаров в вашей корзине.
Заказ по номеру товара
Найти мой заказ
Посмотреть мою корзину
Выезд
- Продукция
- Витамины и добавки
- О витаминах и пищевых добавках
- Дополнения к руководствам
- Рекомендуемые
- Основы
- Бестселлеры
- Новый и переработанный
- Продажа
- Здоровье всего тела
- Летняя распродажа
- Продажа лабораторных тестов
- Сортировать по типу
- Аминокислоты
- АртроМакс
- Восстановление костей
- Книги и СМИ
- Карнитин
- Cognitex
- CoQ10
- Куркумин / Куркума
- Пищеварительные ферменты
- Рыбий жир и омега
- Геропротект
- Гормоны (DHEA)
- Life Extension Mix
- Буква Витамины
- Магний
- Мелатонин
- Минералы
- Мультивитамины
- Пре и пробиотики
- Ресвератрол
- Магазин по Концерну здоровья
- Активный образ жизни и фитнес
- Антивозрастное средство и долголетие
- Здоровье костей
- Здоровье мозга
- Здоровье пищеварительной системы
- Здоровье глаз
- Уровень глюкозы / уровень сахара в крови
- Здоровье сердца
- Гормональный баланс
- Иммунная поддержка
- Управление воспалением
- Здоровье суставов
- Почки, мочевой пузырь, здоровье мочевыводящих путей
- Здоровье печени / Детоксикация
- Поддержка настроения
- Поддержка здоровья и комфорта нервов
- Сексуальное здоровье
- Сон
- Управление стрессом
- Щитовидная железа / надпочечники
- Диета и образ жизни
- Активный образ жизни и фитнес
- Энергетический менеджмент
- Еда и напитки
- Мужское здоровье
- Уход за домашними животными
- Белок
- Контроль веса
- Код оздоровления
- Женское Здоровье
- Красота и личная гигиена
- Уход за кожей Cosmesis
- Витамины и добавки
.